Clear Sky Science · ru
Рекуррентная эволюция множественности лигандсвязывающих доменов тонко настраивает сигнал TGFβ у позвоночных
Как клетки тонко настраивают свои сообщения
С каждой секундой клетки у животных от рыб до людей обмениваются химическими сообщениями, которые подсказывают им, когда расти, заживать или менять свою идентичность. Одна из важнейших систем сигнализации — TGFβ — долгое время считалась собранной из практически неизменных молекулярных деталей. Это исследование показывает, что, скрытые в геномах многих позвоночных, некоторые из этих компонентов тихо переосмысливались, давая клеткам новые способы увеличивать или уменьшать сигнал, а не просто включать или выключать его.
Система сообщений, лежащая в основе построения тела
Сигнализация TGFβ помогает формировать план тела эмбрионов, направляет решения о судьбе клеток и поддерживает гомеостаз тканей во взрослом организме. Сообщение начинается вне клетки, где белковые посланцы, или лиганды, связываются с рецепторами на поверхности клетки. Каждый рецептор имеет наружную «хватку», которая улавливает лиганд, сегмент, проходящий через мембрану, и внутреннюю ферментную область, передающую сигнал внутрь. Когда правильные лиганды связываются, пары рецепторов типов I и II собираются в четырехкомпонентный комплекс и активируют белки SMAD, которые затем попадают в ядро и корректируют активность генов.
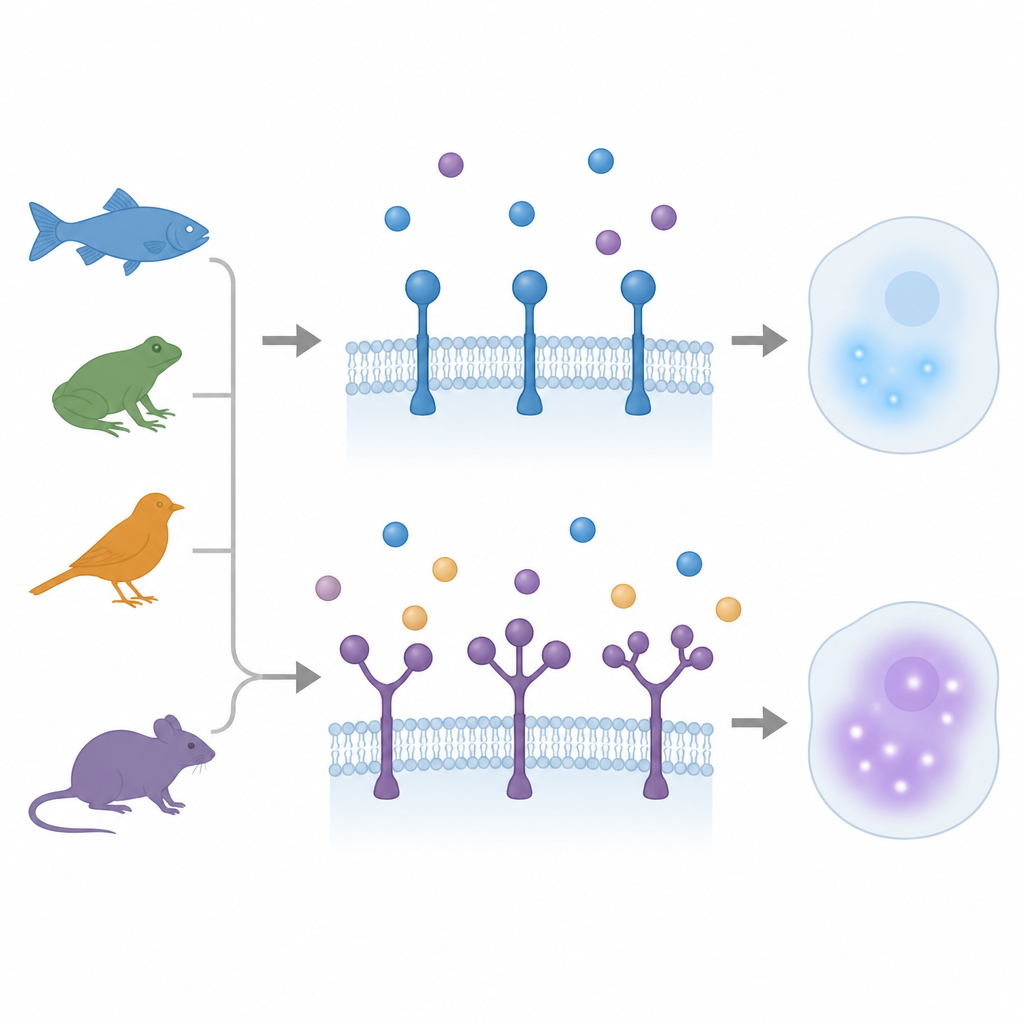
Когда одна «хватка» становится множественной
Годы считалось, что рецепторы семейства TGFβ несут только одну доменную «хватку» для лиганда. Ранее в маленькой рыбе медака обнаружили сюрприз: один рецептор, ACVR1, имел три повторяющихся домена хватки. В новом исследовании авторы просканировали геномы и транскриптомные данные позвоночных в широком эволюционном диапазоне — от лучеперых рыб и латимерий до амфибий, птиц и млекопитающих. Они выявили 12 отдельных случаев, в которых у трех типов рецепторов — ACVR1, BMPR2 и TGFBR2 — эволюционировали дополнительные копии доменов хватки, иногда удваиваясь, иногда утраиваясь. Эти события происходили независимо в разных линиях, что показывает, что природа многократно возвращалась к одному и тому же структурному приему.
Дополнительные «хватки», которые помогают, мешают или делят нагрузку
Команда затем спросила, что на самом деле делают добавленные домены. Используя моделирование структур, докинг‑симуляции и клеточные тесты связывания, они изучили рецепторы с двумя или тремя наружными «хватками». У рыбьих рецепторов BMPR2 с тремя хватками самый внутренний домен, ближайший к мембране, сохранил ключевые контактные точки для лиганда Activin и показал наибольшее предсказанное и измеренное связывание. Более удаленные хватки связывали слабо и действовали как тормоза; их удаление усиливало сигнализацию, хотя у рецептора оставалось меньше физических точек контакта. Похожая картина наблюдалась у нескольких вариантов TGFBR2: внутренний домен выполнял большую часть полезного связывания и активации сигнала, тогда как внешний домен с более быстрыми эволюционными изменениями вел себя скорее как регулируемый демпфер.
Разные виды — разные стратегии настройки
Не все виды использовали дуплицированные домены одинаково. У кур и некоторых млекопитающих, например лошадей, оба домена в TGFBR2 оставались высоко схожими по последовательности и имели почти идентичную поверхность связывания. Любая из хваток могла по‑отдельности обеспечивать сильное связывание лиганда и передачу сигнала, а двухдоменная версия особенно эффективно улавливала лиганд, не теряя выходного ответа. У же в данио‑рерио (зебрафиш), однако, один ген TGFBR2 несет две сильно отличающиеся хватки, а второй, более простой «партнер», содержит единственную хватку. Комплексная версия связывает лиганд, но дает более слабый ответ вниз по потоку, чем ее собрат, и экспрессируется преимущественно в определенных кроветворных и мезодермальных тканях. Переэкспрессия этих рецепторов в эмбрионах вызывала разные эффекты развития, что поддерживает идею о том, что добавление хваток может создавать вариант с низкой активностью, специализированный для тонкого контроля в отдельных типах клеток.
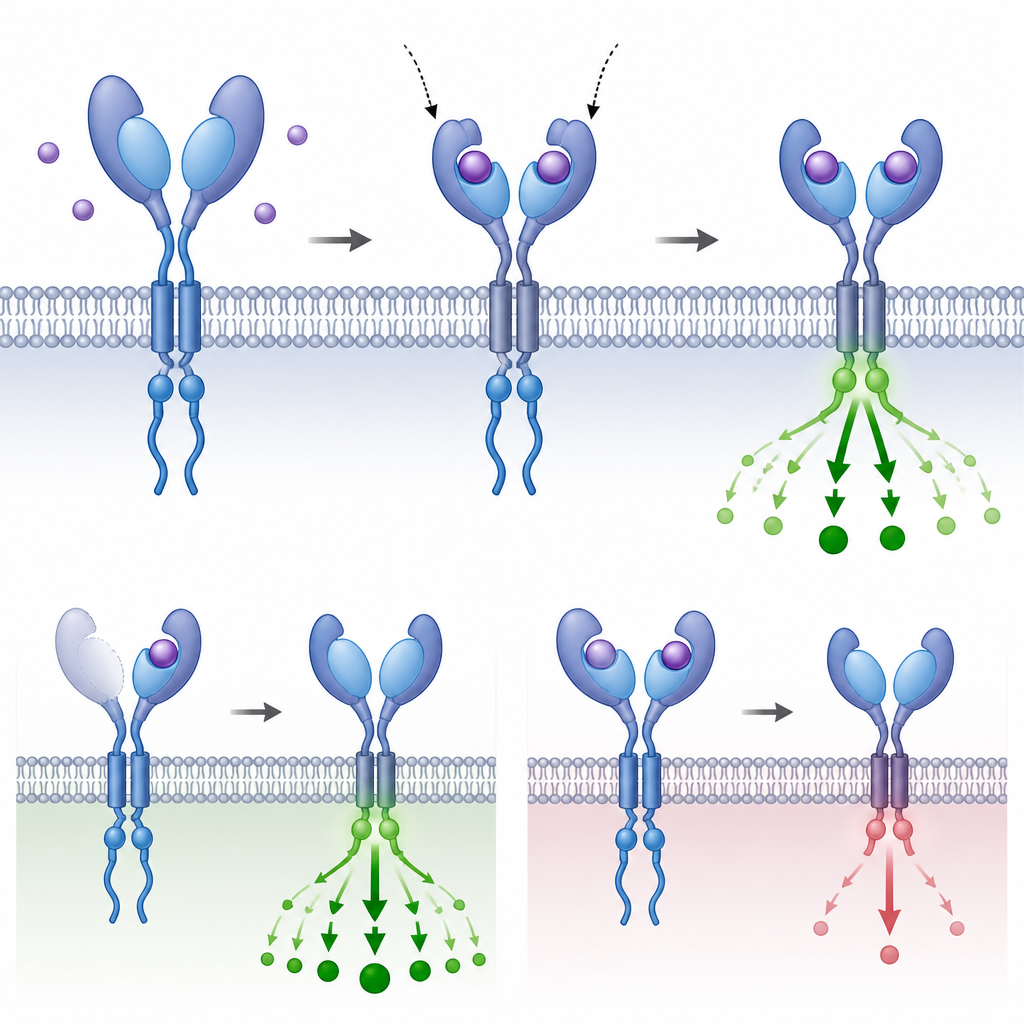
Почему повторение частей важно для эволюции
Прослеживая, где и как эволюционировали эти дополнительные домены хватки и тестируя их поведение в клетках и эмбрионах, авторы показывают, что повторение небольшой части рецептора может изменить то, насколько сильно клетка «слышит» входящее сообщение, не меняя при этом базовой проводки пути. Иногда дополнительные домены усиливают захват лиганда; в других случаях они смягчают или тормозят передачу. Это повторяющееся перестраивание в отдаленных ветвях позвоночных показывает, что дупликация доменов — гибкий эволюционный инструмент, позволяющий организам настраивать глубоко консервативную систему сигнализации под конкретные требования их планов тела и жизненных историй.
Цитирование: Jatzlau, J., Trumpp, M., Kühlwein, J. et al. Recurrent evolution of ligand-binding domain multiplicity fine-tunes TGFβ signaling in vertebrates. Nat Commun 17, 4458 (2026). https://doi.org/10.1038/s41467-026-73340-5
Ключевые слова: сигнализация TGFβ, домены связывания лиганда, эволюция позвоночных, клеточные рецепторы, регуляция сигнала