Clear Sky Science · pl
Wielokrotna ewolucja domen wiążących ligand dopracowuje sygnalizację TGFβ u kręgowców
Jak komórki precyzują swoje komunikaty
Co sekundę komórki zwierząt — od ryb po ludzi — wymieniają się chemicznymi komunikatami, które mówią, kiedy mają rosnąć, goić się lub zmieniać tożsamość. Jednym z najważniejszych systemów przekazywania informacji jest sygnalizacja TGFβ, od dawna uważana za opartą na niemal niezmiennych elementach molekularnych. To badanie pokazuje, że w genomach wielu kręgowców niektóre z tych elementów w ciszy się przeistoczyły, dając komórkom nowe sposoby zwiększania lub tłumienia sygnału, zamiast jedynie włączania go lub wyłączania.
System komunikacyjny u podstaw budowy ciała
Sygnalizacja TGFβ pomaga kształtować plan ciała zarodków, kieruje decyzjami dotyczącymi losu komórek i wspiera równowagę tkanek u dorosłych organizmów. Komunikat zaczyna się na zewnątrz komórki, gdzie białkowe posłańce, czyli ligandy, wiążą się z receptorami na powierzchni komórki. Każdy receptor ma odsłonięty region „uchwytu”, który łapie ligand, odcinek przechodzący przez błonę oraz wewnętrzną domenę enzymatyczną, przekazującą sygnał do wnętrza. Gdy odpowiednie ligandy się przyłączą, pary receptorów typu I i II grupują się w czteroczęściowy kompleks i aktywują białka SMAD, które następnie przemieszczają się do jądra, by dostosować aktywność genów.
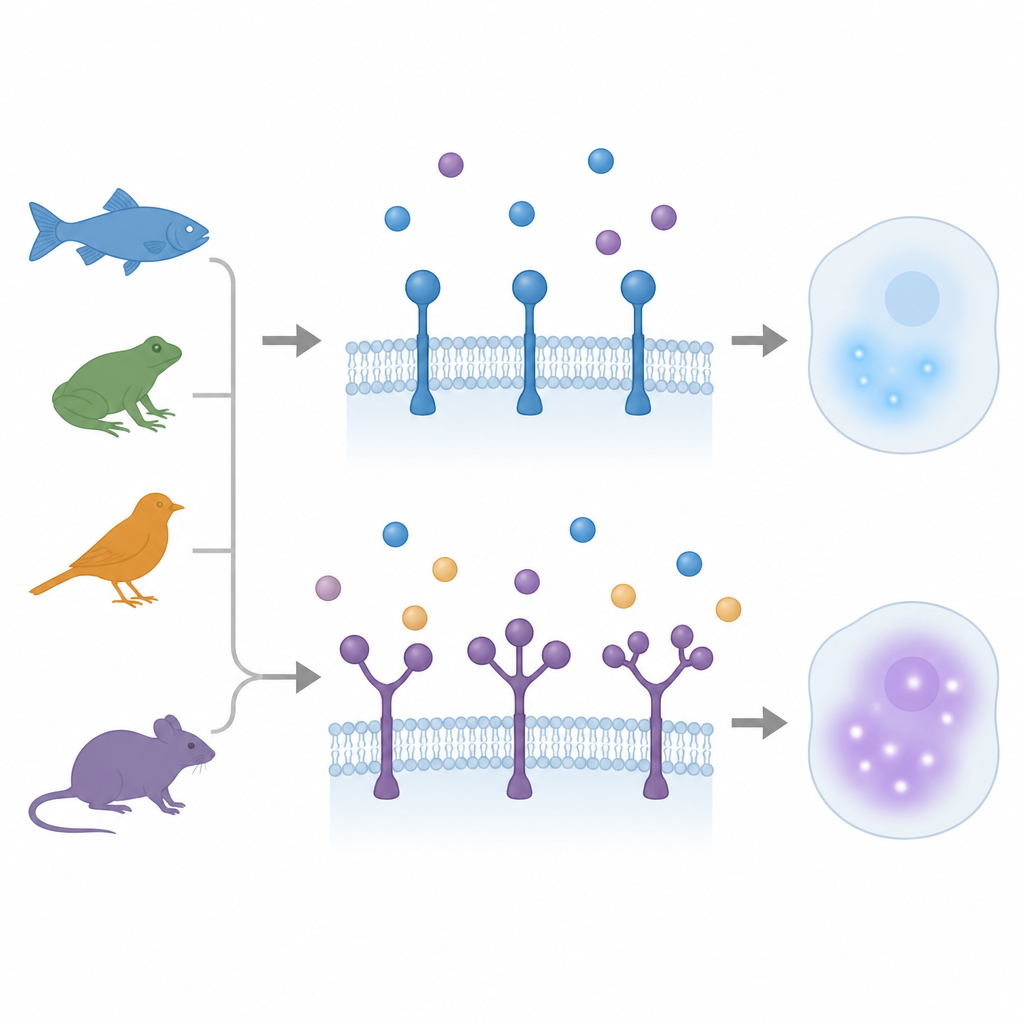
Kiedy jeden uchwyt staje się wieloma
Przez lata uważano, że receptory rodziny TGFβ mają tylko pojedynczy uchwyt wiążący ligand. Wcześniejsze badania na małej rybce medaka odkryły niespodziankę: jeden receptor, ACVR1, miał trzy powtarzające się regiony uchwytu. W nowym badaniu autorzy przeszukali genomy i dane transkryptomiczne kręgowców na szeroką skalę ewolucyjną — od ryb promieniopłetwych i płucopławów po płazy, ptaki i ssaki. Odkryli 12 odrębnych przypadków, w których trzy typy receptorów — ACVR1, BMPR2 i TGFBR2 — nabyły dodatkowe kopie uchwytów, czasem podwajając, a czasem potrajając tę domenę. Zdarzenia te miały miejsce niezależnie w różnych liniach filogenetycznych, co pokazuje, że natura wielokrotnie sięgała po to samo strukturalne rozwiązanie.
Dodatkowe uchwyty, które pomagają, hamują lub dzielą pracę
Zespół zapytał następnie, co te dodane domeny faktycznie robią. Wykorzystując modelowanie struktur, symulacje dokowania i komórkowe testy wiązania, zbadali receptory, których zewnętrzne powierzchnie nosiły dwa lub trzy uchwyty. W rybich receptorach BMPR2 z trzema uchwytami najbliższy błonie uchwyt zachował kluczowe punkty kontaktu z ligandem Activin i wykazywał najsilniejsze przewidywane i zmierzone wiązanie. Dalsze uchwyty wiązały słabo i działały jak hamulce; ich usunięcie zwiększało sygnalizację, mimo że receptor miał mniej fizycznych punktów kontaktu. Podobny wzorzec pojawił się w kilku wariantach TGFBR2: wewnętrzny uchwyt wykonywał większość użytecznego wiązania i sygnalizacji, podczas gdy zewnętrzny uchwyt o szybszych zmianach ewolucyjnych zachowywał się bardziej jak regulowany tłumik.
Różne gatunki, różne strategie strojenia
Nie wszystkie gatunki wykorzystywały zduplikowane domeny w ten sam sposób. U kur i u niektórych ssaków, takich jak konie, oba uchwyty w TGFBR2 pozostały bardzo podobne w sekwencji i miały niemal identyczną powierzchnię wiążącą. Każdy z uchwytów osobno mógł zapewnić silne wiązanie liganda i sygnalizację, a wersja z dwoma uchwytami szczególnie dobrze przechwytywała ligand, nie tracąc przy tym efektu sygnałowego. W zebrafishu natomiast jeden gen TGFBR2 nosi dwa bardzo różne uchwyty, a drugi, prostszy gen partnerski ma pojedynczy uchwyt. Wersja złożona wiąże ligand, ale przekazuje słabszą odpowiedź downstream niż jej krewniak i jest aktywna głównie w określonych tkankach tworzących krew i mezodermie. Przeekspresjonowanie tych receptorów w zarodkach wywołało różne efekty rozwojowe, co potwierdza tezę, że dodane uchwyty mogą tworzyć wariant o niskiej aktywności, wyspecjalizowany do precyzyjnej kontroli w wybranych typach komórek.
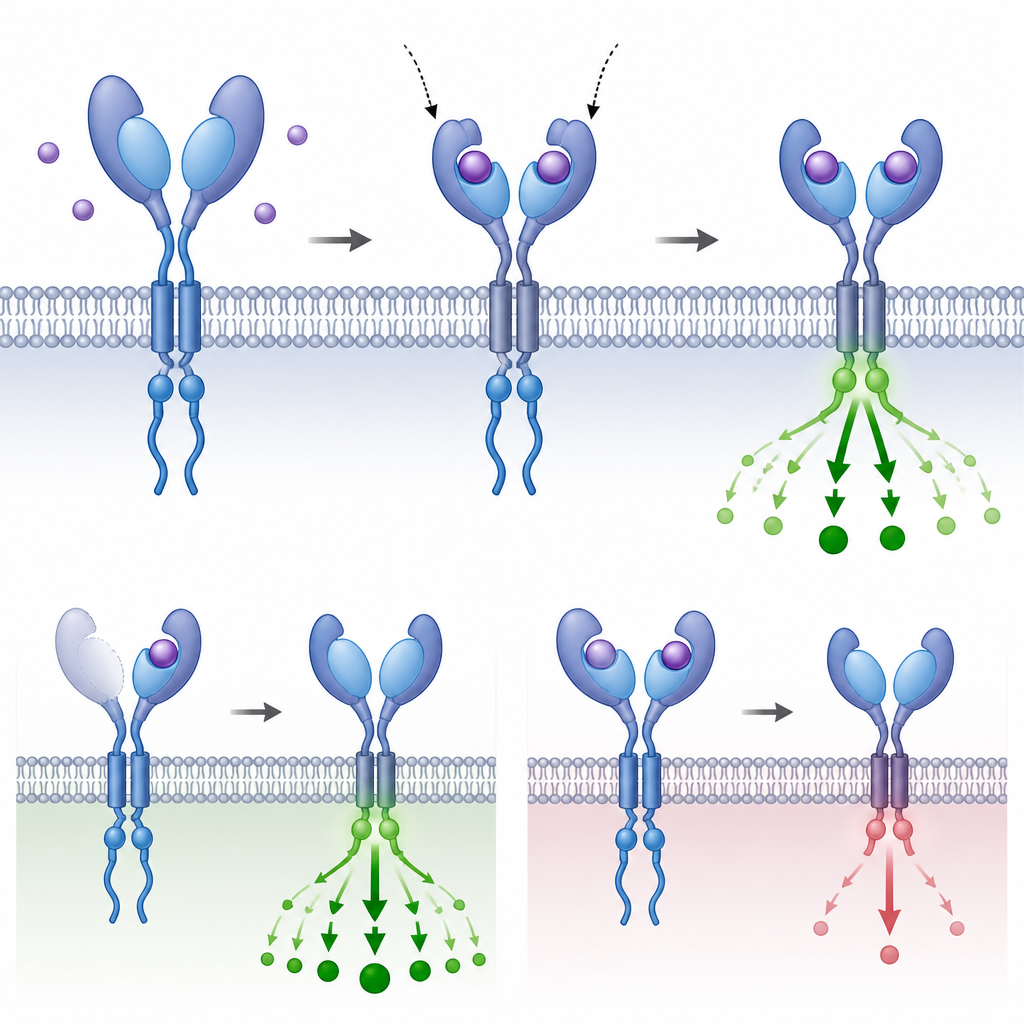
Dlaczego powtarzalne części mają znaczenie dla ewolucji
Śledząc, gdzie i jak te dodatkowe domeny uchwytu ewoluowały oraz testując ich zachowanie w komórkach i zarodkach, autorzy pokazują, że powielenie niewielkiej części receptora może zmienić siłę, z jaką komórka odbiera nadchodzący komunikat, bez modyfikowania podstawowego okablowania szlaku. Czasami dodatkowe domeny zwiększają przechwytywanie liganda; w innych przypadkach buforują lub hamują transmisję. To powtarzające się przeobrażanie w odległych gałęziach kręgowców ujawnia, że duplikacja na poziomie domeny jest elastycznym narzędziem ewolucyjnym, pozwalającym organizmom dostroić głęboko zachowany system sygnalizacyjny do specyficznych wymagań ich planów budowy ciała i historii życiowych.
Cytowanie: Jatzlau, J., Trumpp, M., Kühlwein, J. et al. Recurrent evolution of ligand-binding domain multiplicity fine-tunes TGFβ signaling in vertebrates. Nat Commun 17, 4458 (2026). https://doi.org/10.1038/s41467-026-73340-5
Słowa kluczowe: sygnalizacja TGFβ, domeny wiążące ligand, ewolucja kręgowców, receptory komórkowe, regulacja sygnału