Clear Sky Science · it
L’evoluzione ricorrente della molteplicità dei domini di legame del ligando affina il segnale TGFβ nei vertebrati
Come le cellule affinano i loro messaggi
Ogni secondo, cellule di animali che vanno dai pesci agli esseri umani scambiano messaggi chimici che indicano quando crescere, riparare o cambiare identità. Uno dei sistemi di messaggistica più importanti è la via TGFβ, a lungo ritenuta composta da parti molecolari quasi immutabili. Questo studio dimostra che, nascosti nei genomi di molti vertebrati, alcuni di questi elementi si sono silenziosamente reinventati, offrendo alle cellule nuovi modi per aumentare o diminuire i segnali invece di limitarli a un semplice acceso/spento.
Il sistema di messaggistica al centro della costruzione corporea
La segnalazione TGFβ contribuisce a plasmare il piano corporeo degli embrioni, guida le decisioni sul destino cellulare e sostiene l’equilibrio dei tessuti nell’adulto. Il messaggio ha inizio all’esterno della cellula, dove proteine messaggere, o ligandi, si legano a recettori sulla superficie cellulare. Ogni recettore possiede una regione di “presa” esposta che cattura il ligando, un segmento che attraversa la membrana e una regione enzimatica interna che trasmette il segnale all’interno. Quando i ligandi appropriati si legano, coppie di recettori di tipo I e II si raggruppano in un complesso a quattro parti e attivano le proteine SMAD, che poi migrano al nucleo per modificare l’attività genica.
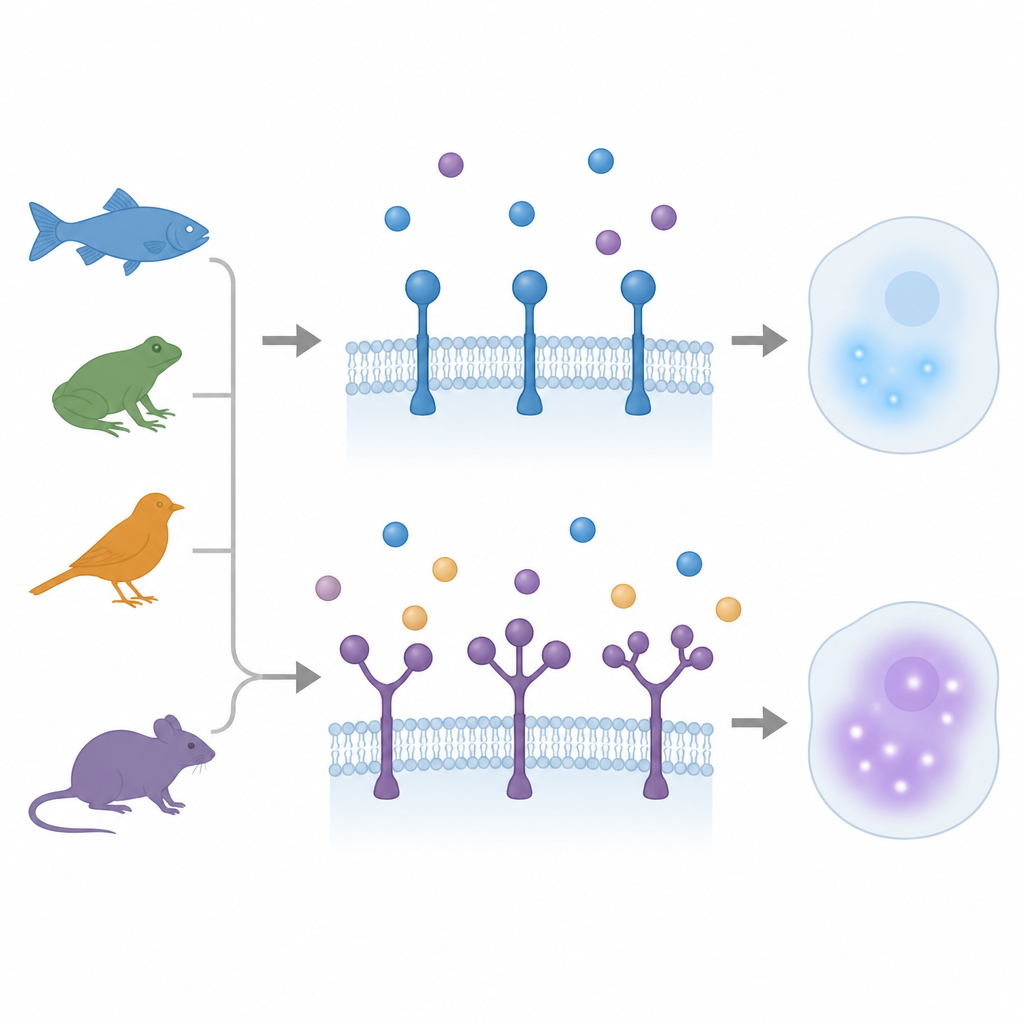
Quando una presa diventa molte
Per anni si è ritenuto che i recettori della famiglia TGFβ portassero una sola presa per il ligando. Lavori precedenti in un piccolo pesce chiamato medaka avevano rivelato una sorpresa: un recettore, ACVR1, presentava tre regioni di presa ripetute. Nel nuovo studio, gli autori hanno scandagliato genomici e dati di trascrittoma dei vertebrati su un ampio spettro evolutivo, dai pesci a pinna raggiata e dai latimerie agli anfibi, uccelli e mammiferi. Hanno scoperto 12 casi separati in cui tre tipi di recettori — ACVR1, BMPR2 e TGFBR2 — hanno evoluto copie aggiuntive di prese, talvolta duplicando e talvolta triplicando il dominio. Questi eventi sono avvenuti in linea indipendente in diversi rami filogenetici, mostrando che la natura è tornata più volte allo stesso espediente strutturale.
Prese aggiuntive che aiutano, ostacolano o condividono il carico
Il gruppo si è allora chiesto quale fosse la funzione concreta di questi domini aggiunti. Utilizzando modellazione strutturale, simulazioni di docking e saggi cellulari di legame, hanno esaminato recettori le cui superfici esterne portavano due o tre prese. Nei recettori BMPR2 dei pesci con tre prese, la presa più interna, più vicina alla membrana, manteneva punti di contatto chiave per il ligando Activin e mostrava il legame previsto e misurato più forte. Le prese più distali si legavano debolmente e fungevano da freni; la loro rimozione aumentava la segnalazione nonostante il recettore avesse meno punti fisici di contatto. Un pattern simile è emerso in diverse varianti di TGFBR2: la presa interna eseguiva la maggior parte del legame e della segnalazione utile, mentre una presa esterna con cambiamenti evolutivi più rapidi si comportava più come un attenuatore regolabile.
Diverse specie, diverse strategie di taratura
Non tutte le specie hanno utilizzato i loro domini duplicati allo stesso modo. Nel pollo e in alcuni mammiferi come i cavalli, entrambe le prese in TGFBR2 sono rimaste molto simili nella sequenza e condividevano una superficie di legame quasi identica. Ciascuna presa da sola poteva sostenere un forte legame col ligando e attivare la segnalazione, e la versione a due prese catturava il ligando in modo particolarmente efficace senza perdere output. Nel zebrafish, invece, un gene TGFBR2 porta due prese molto diverse e un secondo gene partner, più semplice, possiede una singola presa. La versione complessa si lega al ligando ma produce una risposta a valle più debole rispetto al suo omologo ed è espressa principalmente in certi tessuti emopoietici e mesodermici. L’espressione eccessiva di questi recettori negli embrioni ha prodotto effetti di sviluppo distinti, sostenendo l’idea che le prese aggiunte possano creare una variante a segnalazione bassa, specializzata per il controllo fine in tipi cellulari selezionati.
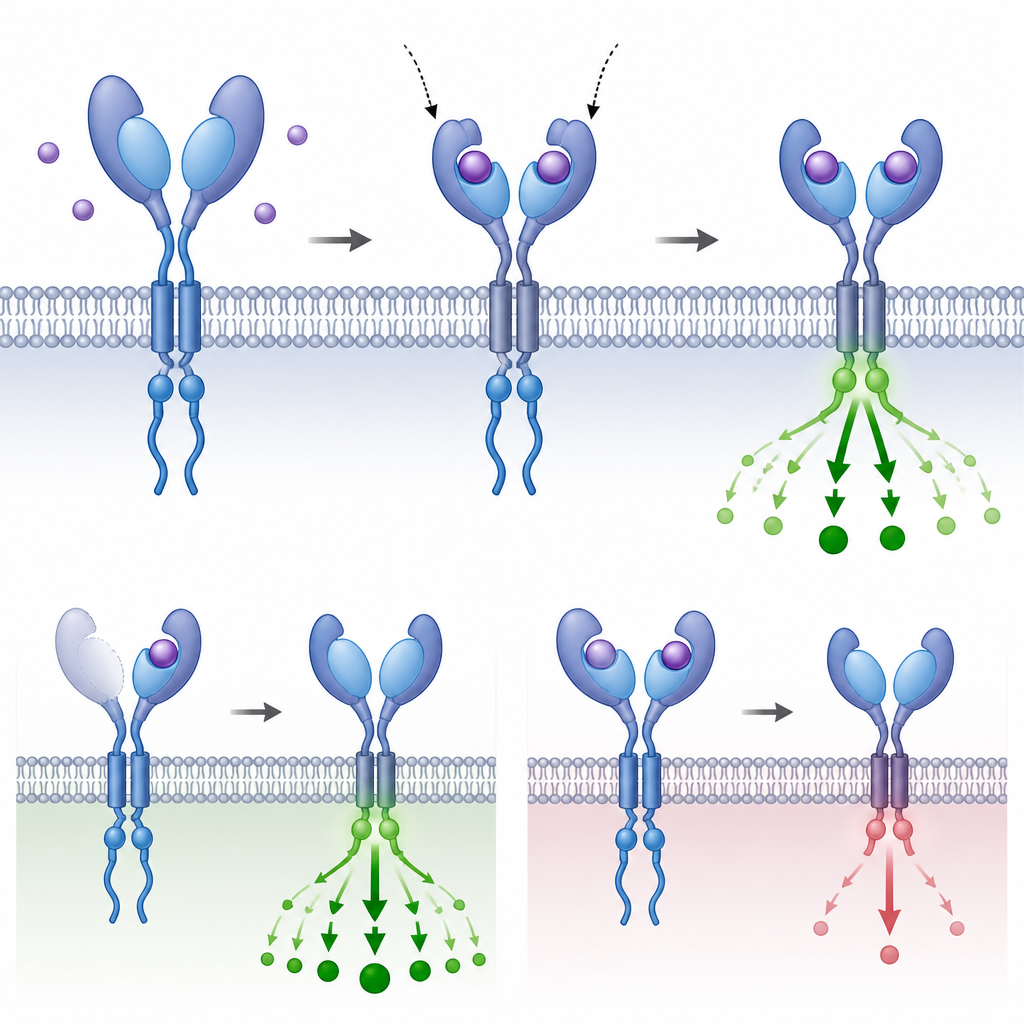
Perché le parti ripetute contano per l’evoluzione
Tracciando dove e come si sono evoluti questi domini di presa aggiuntivi e testandone il comportamento in cellule ed embrioni, gli autori dimostrano che ripetere una piccola parte di un recettore può rimodellare l’intensità con cui una cellula percepisce un segnale in entrata senza cambiare l’architettura di base della via. Talvolta i domini aggiuntivi aumentano la cattura del ligando; in altri casi attenuano o inibiscono la trasmissione. Questo rimodellamento ricorrente in rami vertebrati distanti rivela che la duplicazione a livello di dominio è uno strumento evolutivo flessibile, che permette agli organismi di regolare un sistema di segnalazione profondamente conservato alle esigenze specifiche dei loro piani corporei e delle loro storie di vita.
Citazione: Jatzlau, J., Trumpp, M., Kühlwein, J. et al. Recurrent evolution of ligand-binding domain multiplicity fine-tunes TGFβ signaling in vertebrates. Nat Commun 17, 4458 (2026). https://doi.org/10.1038/s41467-026-73340-5
Parole chiave: segnalazione TGFβ, domini di legame del ligando, evoluzione dei vertebrati, recettori cellulari, regolazione del segnale