Clear Sky Science · en
Recurrent evolution of ligand-binding domain multiplicity fine-tunes TGFβ signaling in vertebrates
How cells fine tune their messages
Every second, cells in animals from fish to humans trade chemical messages that tell them when to grow, heal, or change identity. One of the most important message systems is called TGFβ signaling, long thought to be built from nearly unchanging molecular parts. This study shows that, hidden in the genomes of many vertebrates, some of these parts have quietly reinvented themselves, giving cells new ways to dial signals up or down rather than simply switching them on or off.
The messaging system at the heart of body building
TGFβ signaling helps shape the body plan of embryos, guides cell fate decisions, and supports tissue balance in adulthood. The message starts outside the cell, where protein messengers, or ligands, bind to receptors on the cell surface. Each receptor has an exposed “grip” region that catches the ligand, a segment that crosses the membrane, and an internal enzyme region that relays the signal inward. When the right ligands bind, pairs of type I and type II receptors cluster into a four part complex and activate SMAD proteins, which then travel to the nucleus to adjust gene activity.
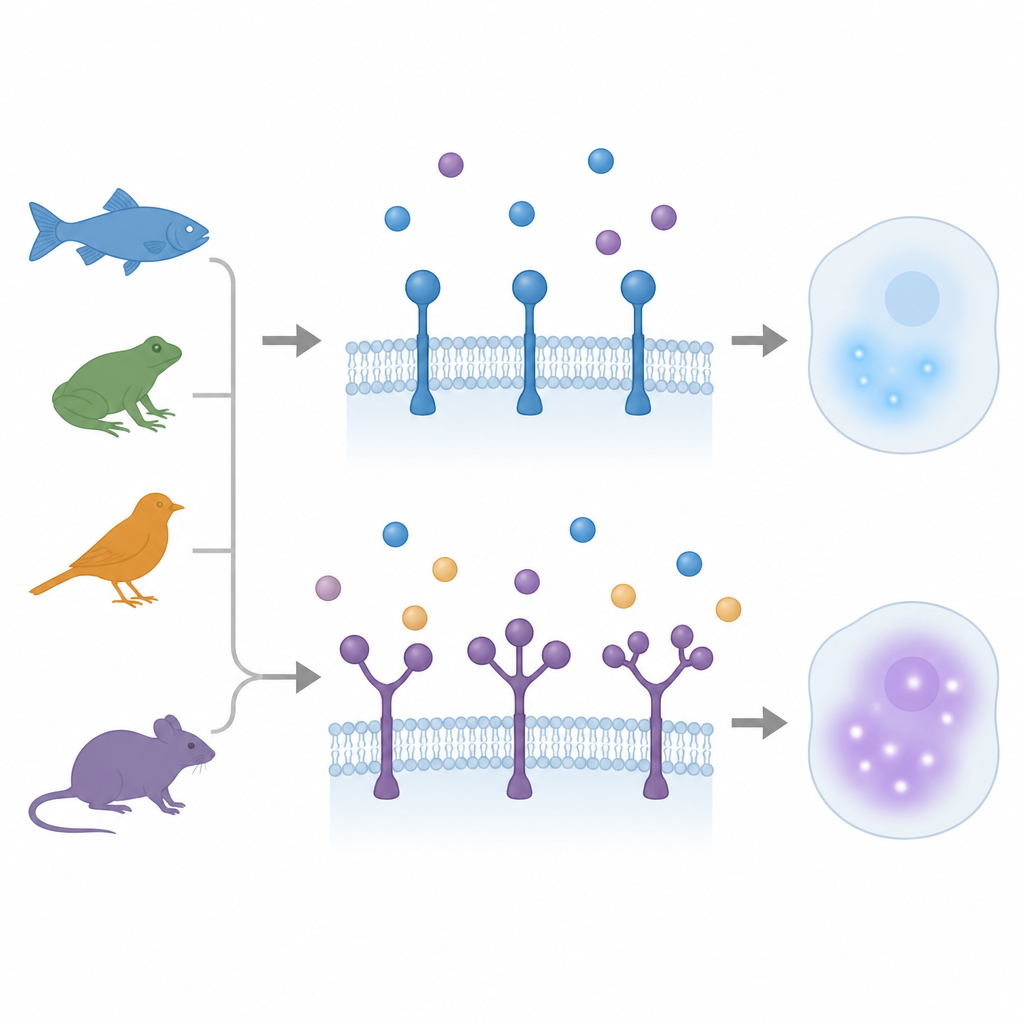
When one grip becomes many
For years, TGFβ family receptors were thought to carry only a single ligand binding grip. Earlier work in a small fish called medaka had uncovered a surprise: one receptor, ACVR1, had three repeated grip regions. In the new study, the authors scanned vertebrate genomes and transcript data across a wide evolutionary range, from ray finned fishes and lungfish to amphibians, birds, and mammals. They uncovered 12 separate cases in which three receptor types ACVR1, BMPR2, and TGFBR2 evolved extra grip copies, sometimes doubling and sometimes tripling the domain. These events occurred independently in different lineages, showing that nature repeatedly returned to the same structural trick.
Extra grips that help, hinder, or share the load
The team then asked what these added domains actually do. Using structure modeling, docking simulations, and cell based binding assays, they examined receptors whose outer surfaces carried two or three grips. In fish BMPR2 receptors with three grips, the innermost grip closest to the membrane retained key contact points for the Activin ligand and showed the strongest predicted and measured binding. The more distant grips bound weakly and acted as brakes; deleting them increased signaling even though the receptor carried fewer physical contact points. A similar pattern appeared in several TGFBR2 variants: the inner grip did most of the useful binding and signaling, while an outer grip with faster evolutionary change behaved more like an adjustable dampener.
Different species, different tuning strategies
Not all species used their duplicated domains the same way. In chickens and some mammals such as horses, both grips in TGFBR2 stayed highly similar in sequence and shared an almost identical binding surface. Either grip alone could support strong ligand binding and signaling, and the two grip version captured ligand especially well without losing output. In zebrafish, however, one TGFBR2 gene carries two very different grips, and a second, simpler partner gene holds a single grip. The complex version binds ligand but gives a weaker downstream response than its sibling, and is turned on mainly in certain blood forming and mesoderm tissues. Overexpressing these receptors in embryos produced distinct developmental effects, supporting the idea that added grips can create a low signaling variant specialized for fine control in select cell types.
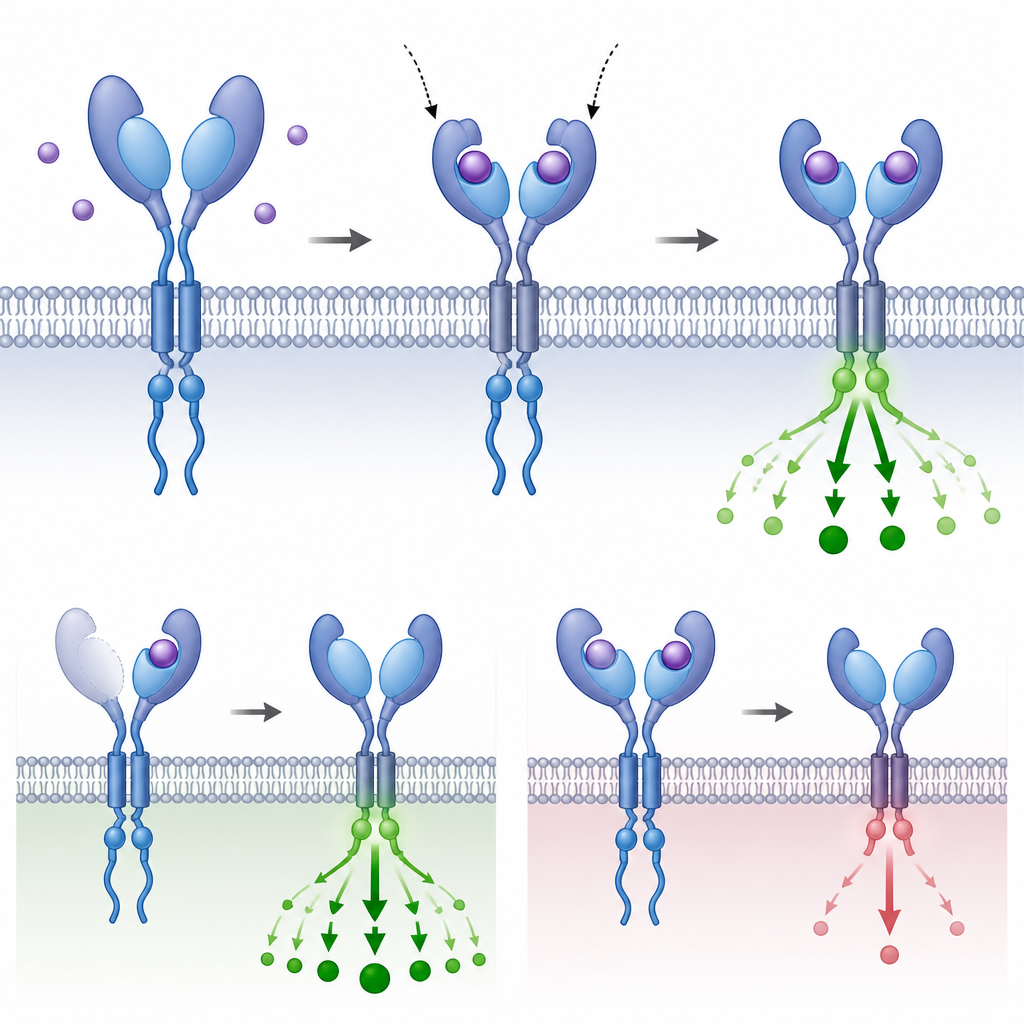
Why repeated parts matter for evolution
By tracing where and how these extra grip domains evolved and testing their behavior in cells and embryos, the authors show that repeating a small part of a receptor can reshape how strongly a cell hears an incoming message without changing the basic wiring of the pathway. Sometimes the extra domains boost ligand capture; in other cases they buffer or inhibit transmission. This recurrent remodeling across distant vertebrate branches reveals that domain level duplication is a flexible evolutionary tool, letting organisms tune a deeply conserved signaling system to the specific demands of their body plans and life histories.
Citation: Jatzlau, J., Trumpp, M., Kühlwein, J. et al. Recurrent evolution of ligand-binding domain multiplicity fine-tunes TGFβ signaling in vertebrates. Nat Commun 17, 4458 (2026). https://doi.org/10.1038/s41467-026-73340-5
Keywords: TGFβ signaling, ligand binding domains, vertebrate evolution, cell receptors, signal regulation