Clear Sky Science · pl
Powstająca elastyczność nabłonków rządzona przez mechanikę powierzchni międzyfazowych i interakcję z podłożem
Jak żywe tkanki się wyginają i marszczą
Nasza skóra, jelita i wiele narządów wyścielają cienkie warstwy komórek zwane nabłonkami. Te żywe arkusze wyginają się, fałdują i marszczą w trakcie rozwoju zarodkowego oraz gdy tkanki dorosłe naprawiają się i adaptują. Artykuł stawia pozornie proste pytanie: czy te tkanki deformują się jak zwykłe materiały sprężyste, takie jak metalowe blachy czy gumowe płachty, czy ich komórkowa natura powoduje, że rządzą nimi inne zasady fizyczne? Budując nową teorię opartą na mechanice na poziomie komórkowym, autorzy pokazują, że nabłonki mogą zachowywać się w zaskakujący sposób, którego nie da się wytłumaczyć za pomocą standardowej podręcznikowej elastyczności.
Komórki jak krople, nie jak lite bloki
Tradycyjne modele traktują warstwę nabłonkową jak płytę stałą, której opór przed zginaniem i rozciąganiem jest równomiernie rozłożony w grubości. Autorzy proponują zamiast tego model, w którym każda komórka przypomina małą kroplę ciekłą: jej wnętrze jest niemal nieściśliwe, a większość sił mechanicznych skupiona jest na powierzchni. Różne części powierzchni komórki – wierzch (skierowany do jamy), spód (stykający się z cienką błoną podstawową i miękkim zrębem) oraz boki (gdzie komórki przylegają do siebie) – mogą mieć różne napięcia powierzchniowe. Napięcia te wynikają z molekularnych silników w korze komórkowej i z białek adhezyjnych. Łącząc wiele takich komórek w łańcuch i dopuszczając do przenoszenia naprężeń tylko przez ich powierzchnie, autorzy wyprowadzają, jak cała tkanka reaguje na kompresję.
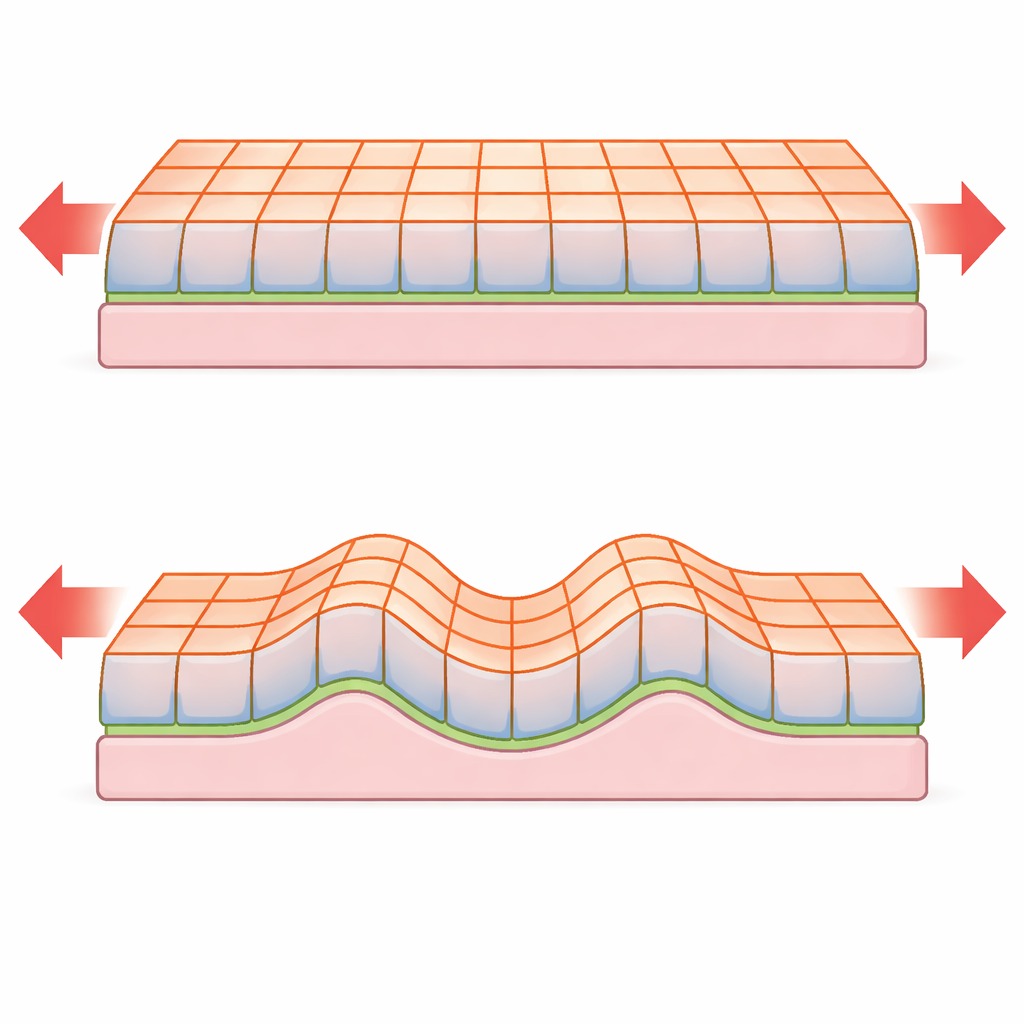
Od reguł na skali komórkowej do zachowania na skali tkanki
Badanie zaczyna się od szczegółowego „modelu wierzchołkowego”, w którym przekrój każdej komórki ma kształt czworokąta, a jego krawędzie niosą określone napięcia powierzchniowe. Tkanka spoczywa na cienkiej, giętkiej błonie podstawowej i grubszej, sprężystej zrębie poniżej niej. Z tego dyskretnego opisu autorzy starannie wyprowadzają teorię ciągłą – zestaw efektywnych parametrów sprężystych opisujących arkusz na większych skalach. W tym uogólnionym podejściu energia deformacji wygląda matematycznie podobnie do energii płyty sprężystej, ale sposób, w jaki kluczowe wielkości zależą od grubości tkanki, jest dramatycznie inny, ponieważ źródłem elastyczności są interfejsy komórkowe, a nie objętość materiału.
Niezwykłe reguły zginania i marszczenia
Kiedy cienka płyta jest ściskana, może albo pozostać płaska i się skrócić, albo wygiąć się poza płaszczyznę i utworzyć fale. Klasyczna teoria płyt przewiduje, że krytyczna siła powodująca zginanie i odstęp między zmarszczkami zależą silnie od grubości płyty. Nowy model pokazuje, że dla arkuszy nabłonkowych rządzonych napięciami powierzchniowymi te skale mogą się zmieniać. Na przykład w niepodpartym monowarstwie nabłonkowym krytyczna siła zginania jest praktycznie niezależna od grubości tkanki, w przeciwieństwie do silnej zależności od grubości przewidywanej dla płyt stałych. Gdy tkanka przylega do błony podstawowej lub osadzona jest na miękkim zrębie, może się marszczyć o długości fali i progu niestabilności, które znów skalują się z grubością inaczej niż w materiałach nieożywionych. Analiza ujawnia również płynne przejście między prostym zginaniem a prawdziwym marszczeniem, gdy wzrasta kontrast między napięciami powierzchniowymi górnej i dolnej strony.
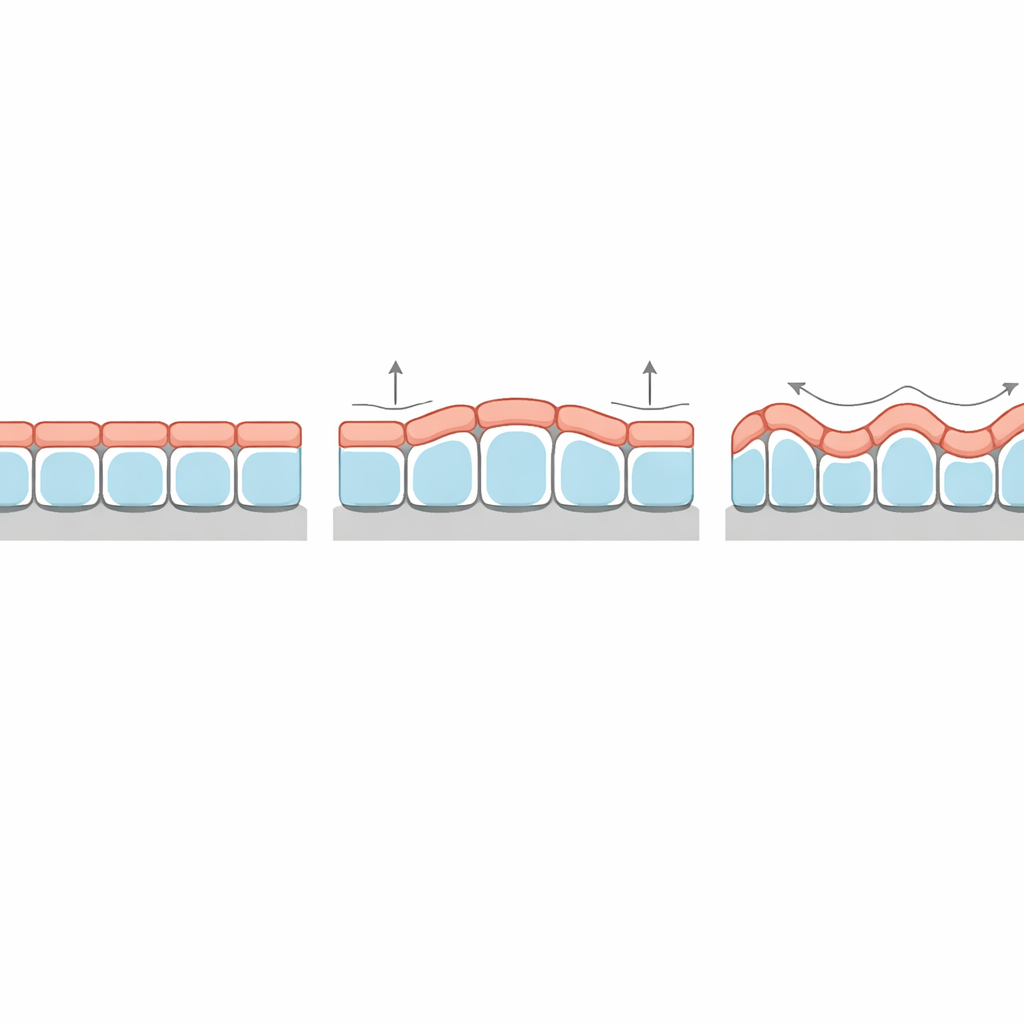
Zagadkowy wzorzec grubości wyjaśniony
Szczególnie uderzającą predykcją jest to, jak grubość tkanki zmienia się wzdłuż grzbietów i dolin jej zmarszczek. W klasycznych bilayerach elastycznych cienka górna warstwa zawsze jest grubsza na grzbietach niż w zagłębieniach, zgodnie z podłożem. Tymczasem niektóre organoidy nabłonkowe i tkanki hodowane na falistych podłożach wykazują odwrotny wzorzec: komórki są wyższe w dolinach niż na szczytach. Nowa teoria pokazuje, że ta „inwersja fazowa” pojawia się naturalnie, gdy napięcie powierzchniowe na wierzchu komórek jest dostatecznie wyższe niż u podstawy. W takich warunkach tkanka minimalizuje całkowitą energię powierzchniową poprzez zmniejszenie silnie rozciągniętej apikalnej powierzchni na grzbietach, skutecznie przesuwając grubość do dolin. Model przewiduje też, jak silnie modulacja grubości występuje i jak zależy to od sztywności błony podstawowej i zrębu.
Dlaczego to ma znaczenie dla biologii i materiałów
Osadzając elastyczność tkanek w mechanice powierzchni komórkowych, praca ta dostarcza fizycznego wyjaśnienia dla kilku obserwacji eksperymentalnych sprzecznych ze standardową teorią płyt, takich jak progi zginania niezależne od grubości i modulacja grubości w przeciwfazie. Sugeruje, że aby zrozumieć, jak narządy fałdują się i marszczą podczas rozwoju, lub jak zaprojektowane tkanki zachowują się na wzorzystych podłożach, trzeba uwzględnić odrębne napięcia na interfejsach komórkowych i ich interakcję z warstwami podtrzymującymi. Poza biologią, ramy te wskazują na szerszą klasę materiałów „międzyfazowych”, których właściwości mechaniczne na dużą skalę wynikają z zaprojektowanych sił powierzchniowych, a nie z sztywności objętościowej, co może inspirować nowe miękkie metamateriały i urządzenia inspirowane biologicznie.
Cytowanie: Andrenšek, U., Krajnc, M. Emergent epithelial elasticity governed by interfacial surface mechanics and substrate interaction. Commun Phys 9, 118 (2026). https://doi.org/10.1038/s42005-026-02547-1
Słowa kluczowe: mechanika nabłonków, zaginanie tkanek, napięcie powierzchniowe, wzory pomarszczeń, morfogeneza