Clear Sky Science · en
Emergent epithelial elasticity governed by interfacial surface mechanics and substrate interaction
How Living Tissues Bend and Wrinkle
Our skin, gut, and many organs are lined by thin layers of cells called epithelia. These living sheets bend, fold, and wrinkle as embryos develop and as adult tissues repair and adapt. This article asks a deceptively simple question: do these tissues deform like ordinary elastic materials, such as metal plates or rubber sheets, or does their cellular nature make them follow different physical rules? By building a new theory grounded in cell-level mechanics, the authors show that epithelia can behave in surprising ways that standard textbook elasticity cannot explain.
Cells as Droplets, Not Solid Blocks
Traditional models treat an epithelial layer as a solid plate whose resistance to bending and stretching is spread uniformly through its thickness. Here, the authors instead model each cell more like a tiny liquid droplet: its interior is nearly incompressible, and most mechanical forces are concentrated at its surface. Different parts of the cell surface – the top (facing a cavity), the bottom (touching a thin basement membrane and soft stroma), and the sides (where cells adhere to each other) – can have different surface tensions. These tensions arise from molecular motors in the cell cortex and from adhesion proteins. By linking many such cells into a chain and letting only their surfaces carry stress, the authors derive how the whole tissue responds when it is compressed.
From Cell-Scale Rules to Tissue-Scale Behavior
The study starts with a detailed “vertex model,” in which each cell cross-section is a four-sided shape whose edges carry specified surface tensions. The tissue rests on a thin, bendable basement membrane and a thicker, elastic stroma beneath it. From this discrete description, the authors carefully derive a continuum theory – a set of effective elastic parameters that describe the sheet on larger scales. In this coarse-grained limit, the energy of deformation looks mathematically similar to that of an elastic plate, but the way key quantities depend on tissue thickness is dramatically different because the origin of elasticity is at cell interfaces rather than in the bulk.
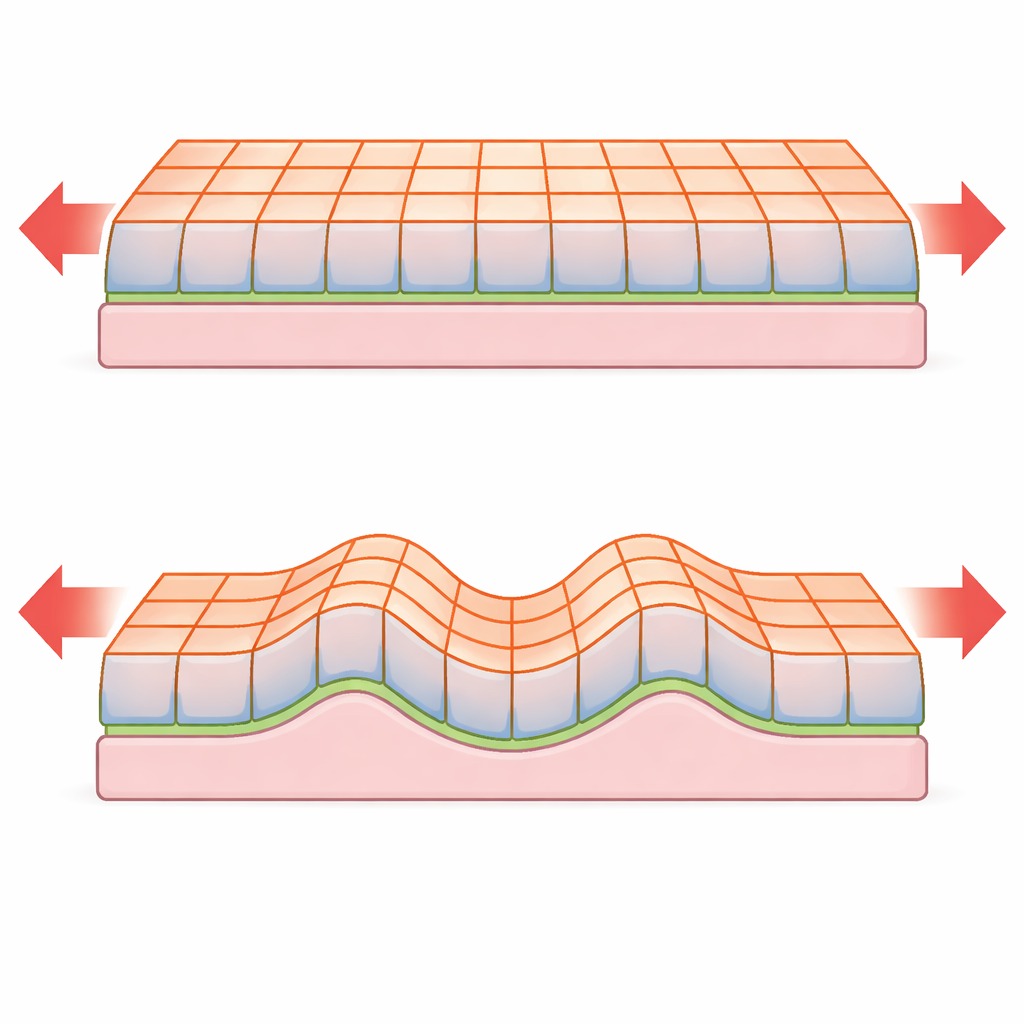
Unusual Buckling and Wrinkling Rules
When a thin plate is compressed, it can either stay flat and shorten, or buckle out of the plane and form waves. Classical plate theory predicts that the critical force for buckling and the spacing between wrinkles depend strongly on the plate’s thickness. The new model shows that for epithelial sheets governed by surface tensions, these scalings can change. For example, in an unsupported epithelial monolayer the critical buckling force is essentially independent of tissue thickness, in contrast to the strong thickness dependence predicted for solid plates. When the tissue is attached to a basement membrane or embedded on a soft stroma, it can wrinkle with a wavelength and instability threshold that again scale differently with thickness than in non-living materials. The analysis also reveals a smooth transition between simple buckling and genuine wrinkling as the contrast between top and bottom surface tensions is increased.
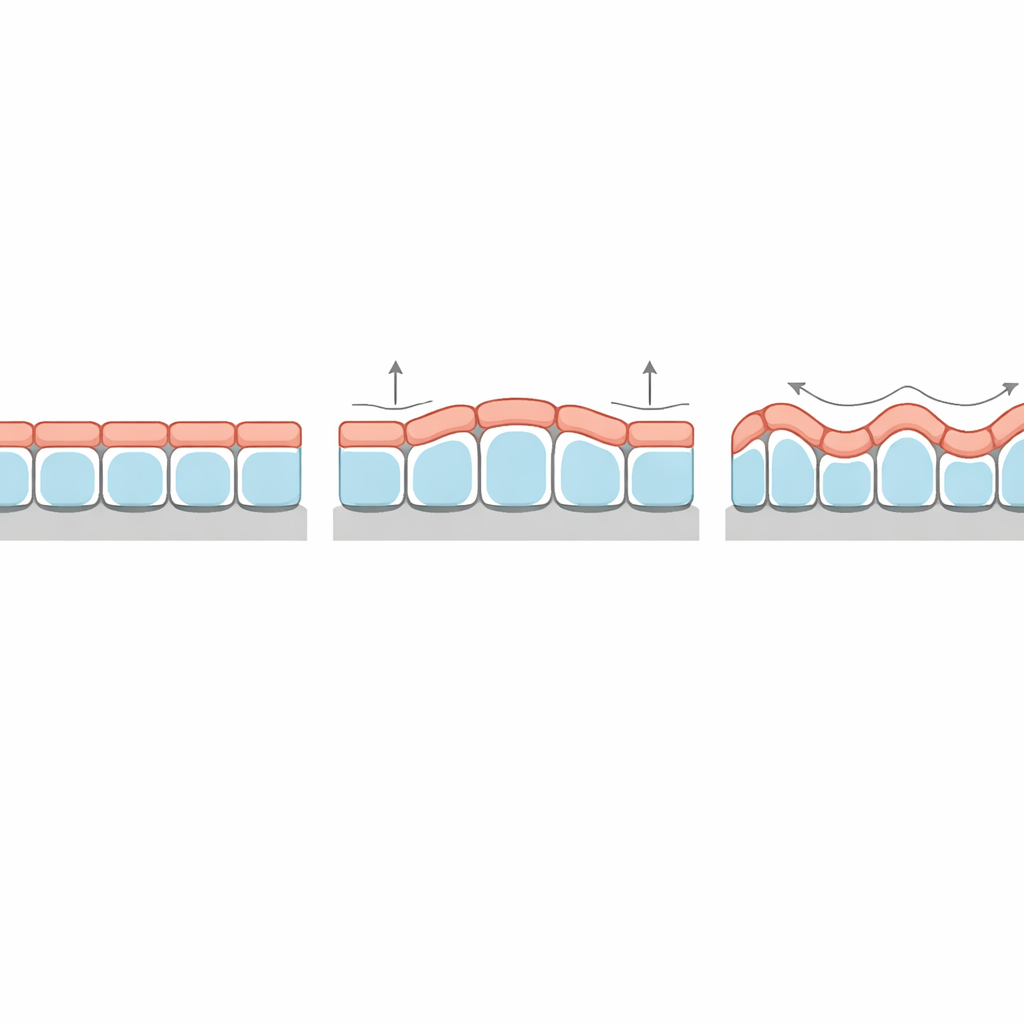
A Puzzling Thickness Pattern Explained
An especially striking prediction concerns how the tissue’s thickness varies along the crests and troughs of its wrinkles. In classic elastic bilayers, the thin top layer is always thicker at the crests than in the grooves, in step with the underlying undulations. Yet some epithelial organoids and tissues grown on wavy substrates show the opposite pattern: cells are taller in the grooves than on the peaks. The new theory shows that this “phase inversion” arises naturally when surface tension at the top of the cells is sufficiently higher than at the bottom. Under those conditions, the tissue minimizes its overall surface energy by reducing the highly tensed apical area at the crests, effectively pushing thickness into the grooves. The model also predicts how strongly thickness modulates and how this depends on the stiffness of the basement membrane and stroma.
Why This Matters for Biology and Materials
By rooting tissue elasticity in cell surface mechanics, this work provides a physical explanation for several experimental observations that conflict with standard plate theory, such as thickness-independent buckling thresholds and anti-phase thickness modulation. It suggests that to understand how organs fold and wrinkle during development, or how engineered tissues behave on patterned substrates, one must account for the distinct tensions at cell interfaces and their interaction with supporting layers. Beyond biology, the framework points toward a broader class of “interfacial” materials whose large-scale mechanical properties emerge from carefully designed surface forces rather than bulk stiffness, offering inspiration for new soft metamaterials and bio-inspired devices.
Citation: Andrenšek, U., Krajnc, M. Emergent epithelial elasticity governed by interfacial surface mechanics and substrate interaction. Commun Phys 9, 118 (2026). https://doi.org/10.1038/s42005-026-02547-1
Keywords: epithelial mechanics, tissue buckling, surface tension, wrinkling patterns, morphogenesis