Clear Sky Science · nl
Opkomende epitheliale elasticiteit bepaald door interfaciale oppervlaktemechanica en substraatsturing
Hoe levende weefsels buigen en kreukelen
Onze huid, darm en vele organen zijn bekleed met dunne cellagen die epithelia worden genoemd. Deze levende vellen buigen, vouwen en kreukelen tijdens de ontwikkeling van embryo’s en wanneer volwassen weefsels herstellen en zich aanpassen. Dit artikel stelt een ogenschijnlijk eenvoudige vraag: vervormen deze weefsels zich als gewone elastische materialen, zoals metalen platen of rubberen folies, of zorgt hun cellulaire aard ervoor dat ze andere fysische regels volgen? Door een nieuwe theorie te bouwen die is gebaseerd op celniveau-mechanica, tonen de auteurs aan dat epithelia zich op verrassende manieren kunnen gedragen die de standaard leerboekelasticiteit niet kan verklaren.
Cellen als druppels, niet als vaste blokken
Traditionele modellen beschouwen een epitheliale laag als een vaste plaat waarvan de weerstand tegen buigen en rekken gelijkmatig over de dikte is verdeeld. Hier modelleren de auteurs elke cel eerder als een kleine vloeibaredruppel: de binnenkant is vrijwel onsamendrukbaar en de meeste mechanische krachten zijn geconcentreerd aan het oppervlak. Verschillende delen van het celoppervlak – de bovenkant (gericht naar een holte), de onderkant (in contact met een dunne basale membraan en zacht stroma) en de zijkanten (waar cellen aan elkaar hechten) – kunnen verschillende oppervlaktespanningen hebben. Deze spanningen ontstaan door moleculaire motoren in de celcortex en door adhesie-eiwitten. Door veel zulke cellen in een keten te verbinden en alleen hun oppervlakken spanningen te laten dragen, leiden de auteurs af hoe het gehele weefsel reageert wanneer het wordt samengedrukt.
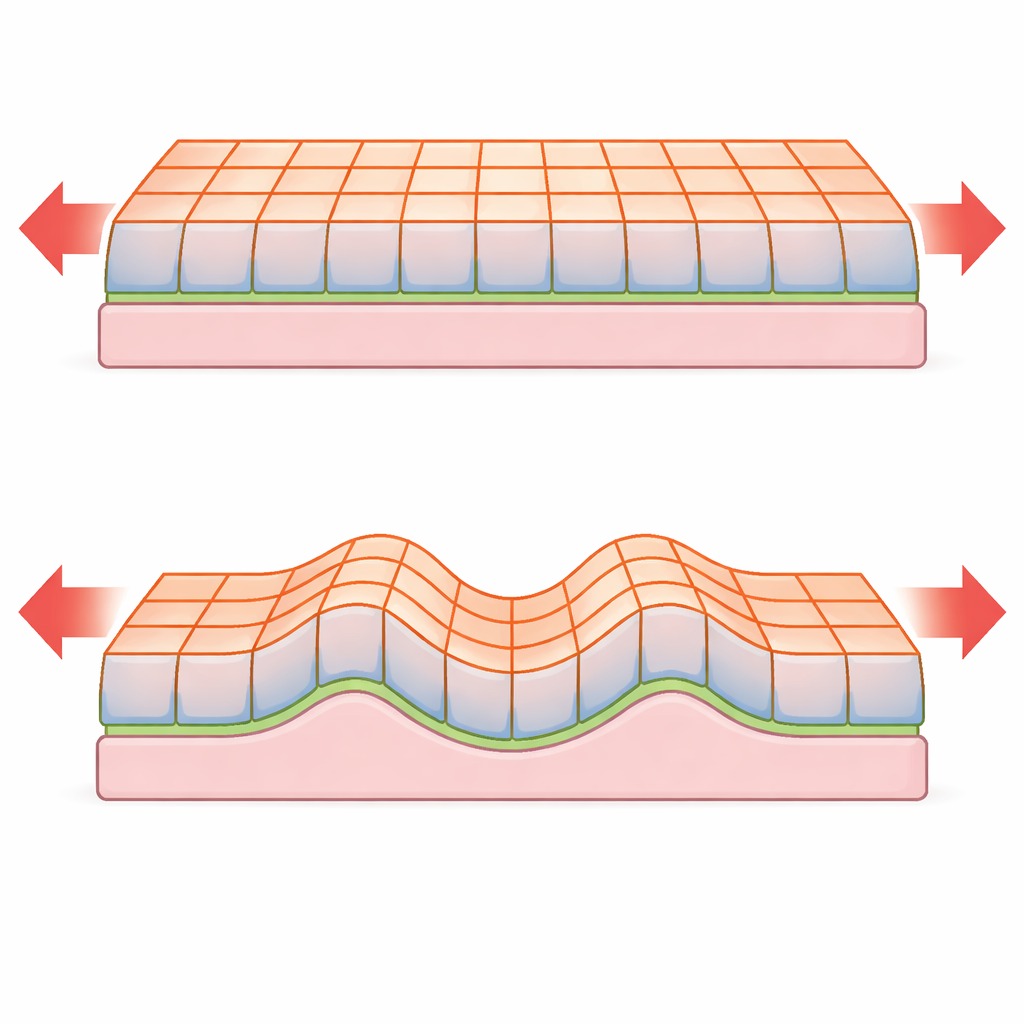
Van celregel naar weefselgedrag
De studie begint met een gedetailleerd “vertexmodel”, waarin elk celdoorsnede een vierzijdige vorm is waarvan de randen opgegeven oppervlaktespanningen dragen. Het weefsel rust op een dun, buigzaam basaal membraan en een dikkere, elastische stroma eronder. Vanuit deze discrete beschrijving leiden de auteurs zorgvuldig een continuümtheorie af — een reeks effectieve elastische parameters die het vel op grotere schalen beschrijven. In deze grofmazige limiet lijkt de vervormingsenergie wiskundig op die van een elastische plaat, maar de manier waarop sleutelgrootheden afhankelijk zijn van weefseldikte is dramatisch anders omdat de oorsprong van elasticiteit in de celinterfaces ligt in plaats van in het bulk.
Ongebruikelijke buckling- en kreukelregels
Wanneer een dunne plaat wordt samengedrukt, kan die ofwel vlak blijven en korter worden, of uit het vlak uitbuigen en golven vormen. De klassieke plaattheorie voorspelt dat de kritische kracht voor buckling en de afstand tussen kreukels sterk afhangen van de dikte van de plaat. Het nieuwe model laat zien dat voor epitheliale vellen die door oppervlaktespanningen worden beheerst, deze schaleringen kunnen veranderen. Bijvoorbeeld, in een onondersteund epitheliaal monolayer is de kritische bucklingkracht in wezen onafhankelijk van de weefseldikte, in tegenstelling tot de sterke dikteafhankelijkheid die voor vaste platen wordt voorspeld. Wanneer het weefsel is bevestigd aan een basaal membraan of ingebed op een zacht stroma, kan het kreukelen met een golflengte en instabiliteitsdrempel die opnieuw anders schalen met de dikte dan bij niet-levende materialen. De analyse onthult ook een vloeiende overgang tussen eenvoudige buckling en echte kreukeling naarmate het contrast tussen boven- en onderoppervlaktespanningen toeneemt.
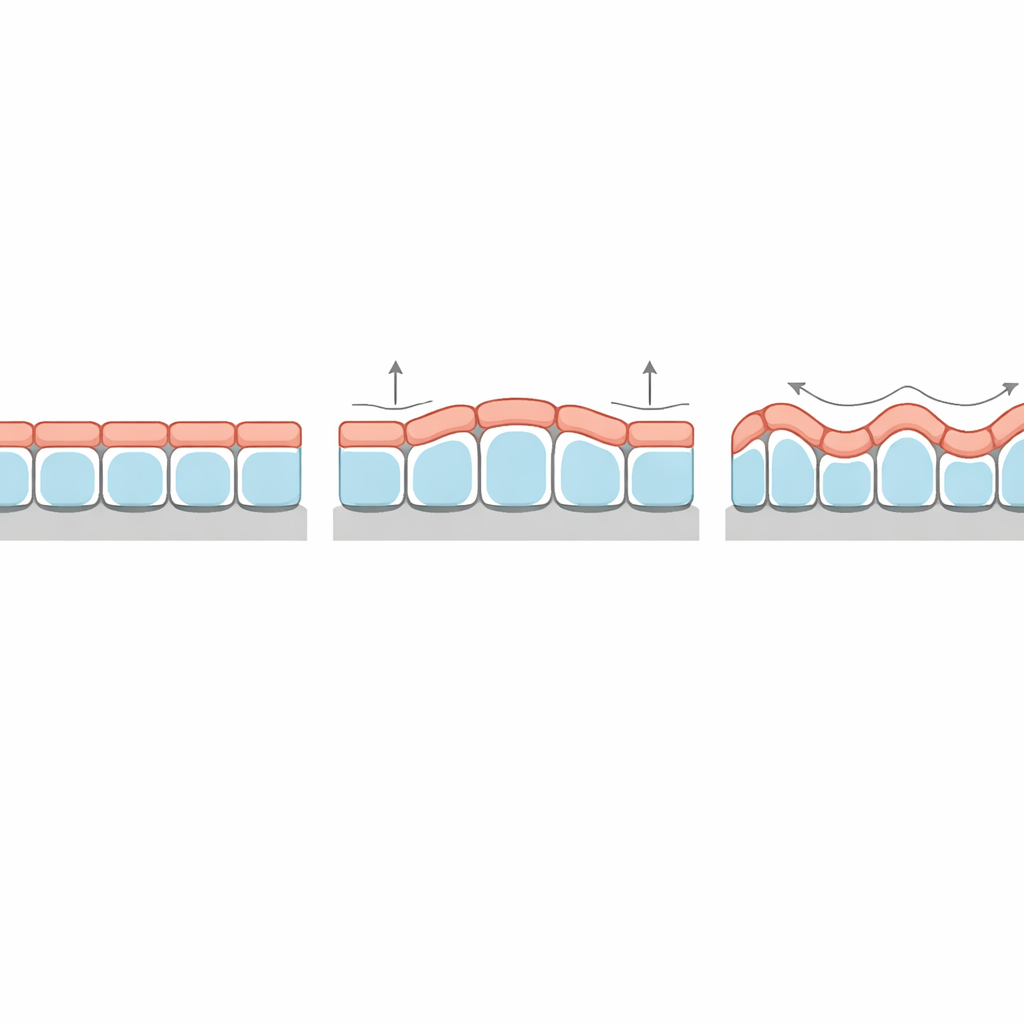
Een raadselachtig diktepatroon verklaard
Een bijzonder opvallende voorspelling betreft hoe de dikte van het weefsel varieert langs de kammen en dalen van zijn kreukels. In klassieke elastische bilagen is de dunne bovenlaag altijd dikker bij de toppen dan in de groeven, in overeenstemming met de onderliggende undulaties. Toch laten sommige epitheliale organoïden en weefsels die op gegolfde substraten zijn gekweekt het omgekeerde patroon zien: cellen zijn hoger in de groeven dan op de pieken. De nieuwe theorie toont dat deze “fase-inversie” vanzelf voorkomt wanneer de oppervlaktespanning aan de bovenkant van de cellen ruim hoger is dan aan de onderkant. Onder die omstandigheden minimaliseert het weefsel zijn totale oppervlakenergie door het sterk gespannen apicale oppervlak bij de toppen te verkleinen, waardoor de dikte effectief naar de groeven wordt gedrukt. Het model voorspelt ook hoe sterk de diktemodulatie is en hoe dit afhangt van de stijfheid van het basale membraan en het stroma.
Waarom dit belangrijk is voor biologie en materialen
Door de elasticiteit van weefsel te funderen in celoppervlaktemechanica, levert dit werk een fysieke verklaring voor verschillende experimentele waarnemingen die in strijd zijn met de standaard plaattheorie, zoals dikte-onafhankelijke bucklingdrempels en anti-fase diktemodulatie. Het suggereert dat om te begrijpen hoe organen vouwen en kreukelen tijdens de ontwikkeling, of hoe gefabriceerde weefsels zich gedragen op gepatroneerde substraten, men rekening moet houden met de onderscheiden spanningen bij celinterfaces en hun interactie met ondersteunende lagen. Buiten de biologie wijst het kader op een bredere klasse van “interfaciale” materialen waarvan de macroscopische mechanische eigenschappen voortkomen uit zorgvuldig ontworpen oppervlaktekrachten in plaats van bulksteifheid, en biedt het inspiratie voor nieuwe zachte metamaterialen en bio-geïnspireerde apparaten.
Bronvermelding: Andrenšek, U., Krajnc, M. Emergent epithelial elasticity governed by interfacial surface mechanics and substrate interaction. Commun Phys 9, 118 (2026). https://doi.org/10.1038/s42005-026-02547-1
Trefwoorden: epitheliale mechanica, weefselbuckling, oppervlaktespanning, kreukelpatronen, morphogenese