Clear Sky Science · es
Elasticidad epitelial emergente gobernada por la mecánica de las superficies interfaciales y la interacción con el sustrato
Cómo se doblan y arrugan los tejidos vivos
Nuestra piel, el intestino y muchos órganos están recubiertos por capas delgadas de células llamadas epitelios. Estas láminas vivas se doblan, pliegan y arrugan durante el desarrollo embrionario y cuando los tejidos adultos se reparan o adaptan. Este artículo plantea una pregunta aparentemente sencilla: ¿se deforman estos tejidos como materiales elásticos ordinarios, como placas metálicas o láminas de caucho, o su naturaleza celular les hace seguir reglas físicas diferentes? Al construir una nueva teoría fundamentada en la mecánica a escala celular, los autores muestran que los epitelios pueden comportarse de formas sorprendentes que la elasticidad de libro de texto no puede explicar.
Las células como gotas, no como bloques sólidos
Los modelos tradicionales tratan la capa epitelial como una placa sólida cuya resistencia al plegado y al estiramiento está distribuida uniformemente a lo largo de su espesor. Aquí, los autores modelan cada célula más bien como una pequeña gota líquida: su interior es casi incompresible y la mayoría de las fuerzas mecánicas se concentran en su superficie. Diferentes partes de la superficie celular —la parte apical (hacia una cavidad), la basal (en contacto con una delgada membrana basal y un estroma blando) y las laterales (donde las células se adhieren entre sí)— pueden presentar tensiones superficiales distintas. Estas tensiones provienen de motores moleculares en el córtex celular y de proteínas de adhesión. Al enlazar muchas de estas células en cadena y dejar que solo sus superficies soporten esfuerzos, los autores deducen cómo responde todo el tejido cuando se lo comprime.
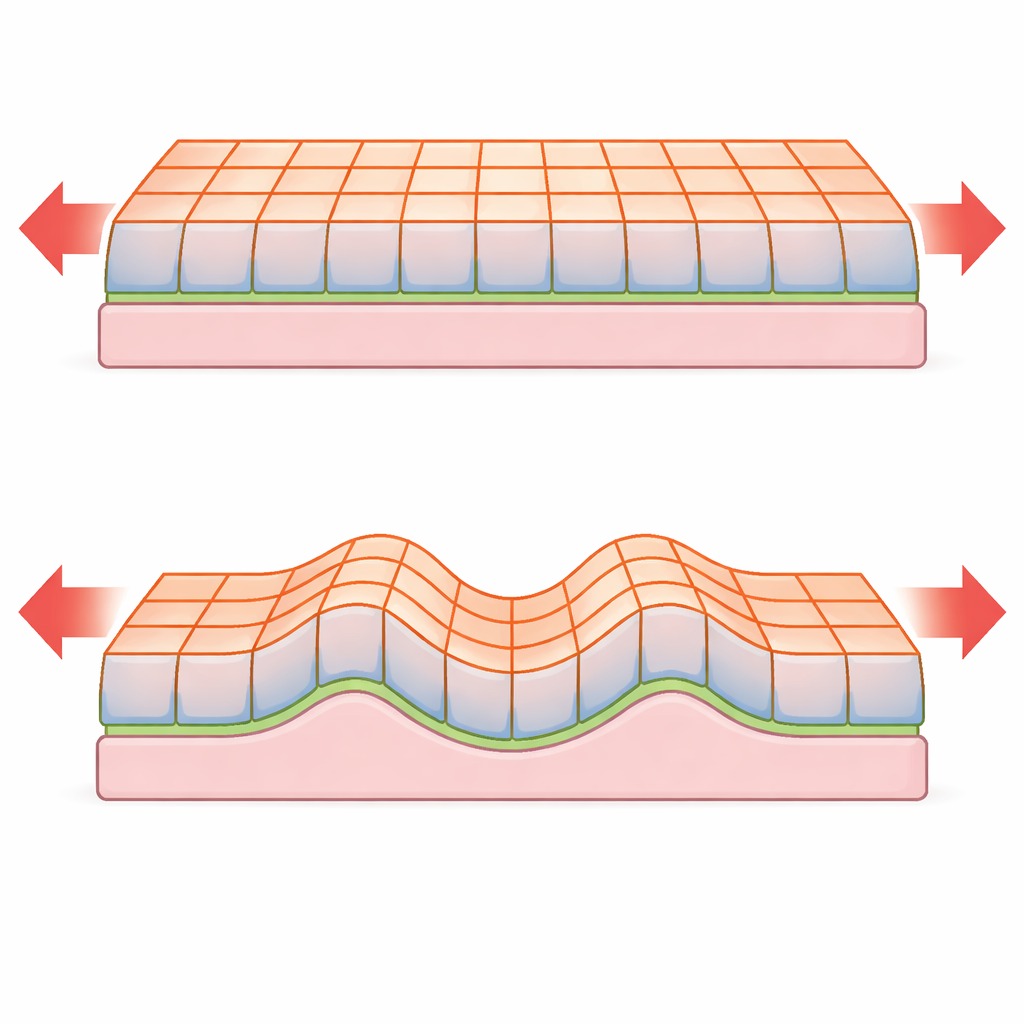
De las reglas a escala celular al comportamiento a escala tisular
El estudio parte de un detallado “modelo de vértices”, en el que la sección transversal de cada célula tiene cuatro lados cuyos bordes soportan tensiones superficiales especificadas. El tejido reposa sobre una membrana basal delgada y flexible y un estroma elástico más grueso debajo. A partir de esta descripción discreta, los autores derivan cuidadosamente una teoría continua: un conjunto de parámetros elásticos efectivos que describen la lámina a escalas mayores. En este límite de corte fino, la energía de deformación se parece matemáticamente a la de una placa elástica, pero la forma en que las cantidades clave dependen del espesor del tejido es drásticamente diferente porque el origen de la elasticidad está en las interfaces celulares y no en el volumen.
Reglas inusuales para pandeo y arrugado
Cuando una placa delgada se comprime, puede permanecer plana y acortarse, o puede salir del plano y formar ondulaciones. La teoría clásica de placas predice que la fuerza crítica para el pandeo y la separación entre arrugas dependen fuertemente del espesor de la placa. El nuevo modelo muestra que, para láminas epiteliales gobernadas por tensiones superficiales, estas escalas pueden cambiar. Por ejemplo, en una monocapa epitelial no soportada, la fuerza crítica de pandeo es esencialmente independiente del espesor del tejido, en contraste con la fuerte dependencia del espesor prevista para placas sólidas. Cuando el tejido está adherido a una membrana basal o apoyado sobre un estroma blando, puede arrugarse con una longitud de onda y un umbral de inestabilidad que, de nuevo, escalan de forma distinta con el espesor respecto a los materiales no vivos. El análisis también revela una transición suave entre el pandeo simple y el arrugado genuino a medida que aumenta el contraste entre las tensiones superficiales apical y basal.
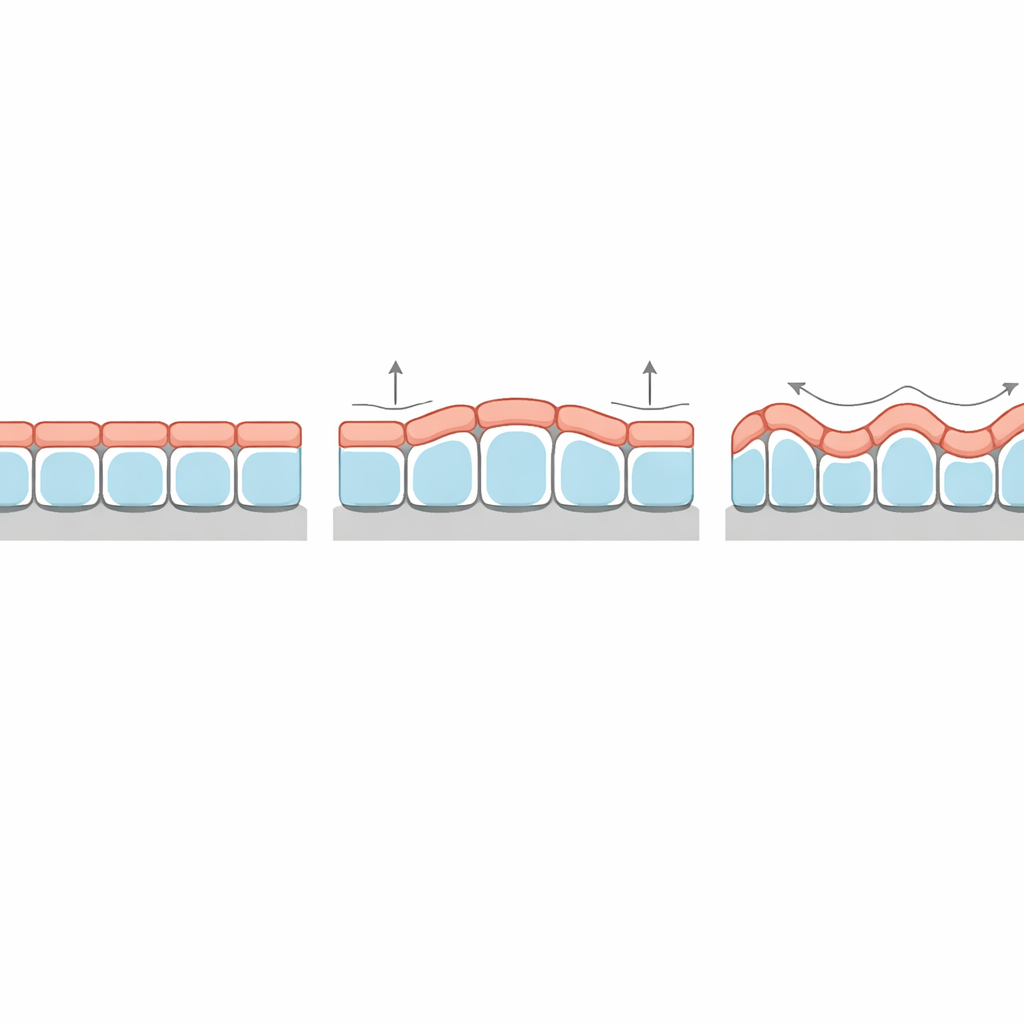
Un patrón de espesor desconcertante explicado
Una predicción especialmente llamativa se refiere a cómo varía el espesor del tejido a lo largo de las crestas y los valles de sus arrugas. En bicapas elásticas clásicas, la capa superior delgada siempre es más gruesa en las crestas que en las hondonadas, siguiendo las ondulaciones subyacentes. Sin embargo, algunos organoides epiteliales y tejidos cultivados sobre sustratos ondulados muestran el patrón opuesto: las células son más altas en las hondonadas que en los picos. La nueva teoría muestra que esta “inversión de fase” surge de forma natural cuando la tensión superficial en la parte apical de las células es suficientemente mayor que en la basal. Bajo esas condiciones, el tejido minimiza su energía superficial global reduciendo el área apical altamente tensa en las crestas, lo que efectivamente desplaza el espesor hacia las hondonadas. El modelo también predice con qué intensidad se modula el espesor y cómo esto depende de la rigidez de la membrana basal y del estroma.
Por qué esto importa para la biología y los materiales
Al arraigar la elasticidad tisular en la mecánica de las superficies celulares, este trabajo ofrece una explicación física para varias observaciones experimentales que chocan con la teoría clásica de placas, como umbrales de pandeo independientes del espesor y modulaciones de espesor en anti-fase. Sugiere que, para comprender cómo se pliegan y arrugan los órganos durante el desarrollo o cómo se comportan los tejidos diseñados en sustratos con patrón, es necesario tener en cuenta las tensiones distintas en las interfaces celulares y su interacción con las capas de soporte. Más allá de la biología, el marco apunta hacia una clase más amplia de materiales “interfaciales” cuyas propiedades mecánicas a gran escala emergen de fuerzas superficiales diseñadas cuidadosamente en lugar de la rigidez del volumen, ofreciendo inspiración para nuevos metamateriales blandos y dispositivos bioinspirados.
Cita: Andrenšek, U., Krajnc, M. Emergent epithelial elasticity governed by interfacial surface mechanics and substrate interaction. Commun Phys 9, 118 (2026). https://doi.org/10.1038/s42005-026-02547-1
Palabras clave: mecánica epitelial, pandeo de tejidos, tensión superficial, patrones de arrugado, morfogénesis