Clear Sky Science · pl
Profilowanie transkrypcyjne i funkcjonalne neuronów z receptorami muskarynowymi w bocznej korze przedczołowej i przedniej korze obręczy u naczelnych
Jak przekaźnik mózgowy kształtuje myślenie i uczucia
W każdej chwili mózg równoważy to, co myślisz, z tym, co czujesz. Dwa kluczowe ośrodki tego procesu to boczna kora przedczołowa, wspomagająca planowanie i pamięć operacyjną, oraz przednia kora obręczy, monitorująca motywację, konflikt i ból. W tym badaniu zadano z pozoru proste pytanie: w jaki sposób ten sam sygnał chemiczny — acetylocholina — dostraja te dwa regiony w różny sposób, aby wspierać uczenie się, elastyczność i kontrolę emocji? 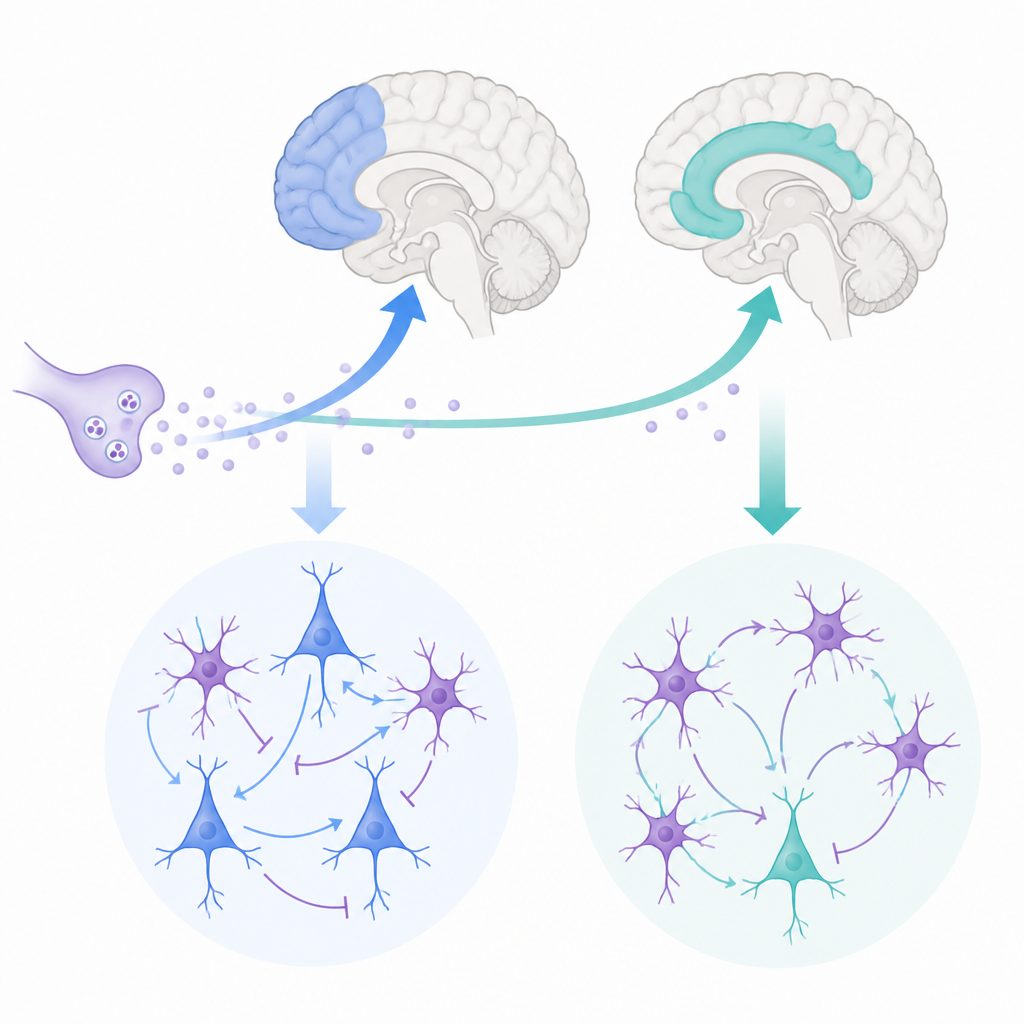
Bliższe spojrzenie na dwa ośrodki myślenia
Naukowcy skupili się na dwóch obszarach płata czołowego makaków rezus, które są silnie podobne do ludzkich struktur. Boczną korę przedczołową angażujemy, gdy utrzymujemy informacje w pamięci i podejmujemy decyzje, podczas gdy przednia kora obręczy wiąże działania z nagrodami, błędami i emocjami. Oba regiony otrzymują sygnały od acetylocholiny — związku uwalnianego przez głębokie ośrodki mózgu, znanego z wpływu na uwagę i pamięć. Jednocześnie przednia kora obręczy otrzymuje gęstsze dopływy acetylocholiny niż boczna kora przedczołowa, co sugeruje, że ten przekaźnik może kształtować ich aktywność na różne sposoby.
Odczytywanie genetycznych odcisków komórek mózgowych
Aby sprawdzić, jak pojedyncze komórki reagują na acetylocholinę, zespół zastosował sekwencjonowanie RNA z pojedynczych jąder komórkowych, które pozwala odczytać, które geny są aktywne w tysiącach komórek jednocześnie. Skoncentrowano się na receptorach muskarynowych — głównych „wyłącznikach” wrażliwych na acetylocholinę na neuronach kory — kodowanych przez cztery geny CHRM1–CHRM4. Ku zaskoczeniu odkryto, że gen CHRM3 był najszerzej eksprymowany w obu regionach, obecny w ponad połowie komórek, choć wcześniejsze badania białkowe sugerowały większe występowanie receptora m1, kodowanego przez CHRM1. Większość neuronów pobudzających i hamujących nosiła CHRM3, często razem z CHRM1, podczas gdy CHRM2 wykazywał bardziej selektywny wzór, szczególnie w głębszych warstwach komórek pobudzających i w istotnej grupie szybko wyładowujących neuronów hamujących.
Kiedy wiadomości genowe i białka nie pasują do siebie
Aby wyjaśnić rozbieżność między RNA a białkiem, naukowcy połączyli fluorescencyjne znakowanie białek receptorowych z detekcją ich RNA in situ w fragmentach mózgu. Potwierdzili, że białko m1 było ogólnie silniejsze niż m3, ale odkryli, że RNA CHRM3 częściej lokalizowało się zarówno w jądrze, jak i w otaczającym cytoplazmie, podczas gdy RNA CHRM1 występowało głównie w cytoplazmie. To sugeruje, że przekazy CHRM3 mogą być zatrzymywane w jądrze lub wolniej przetwarzane na białko, co pomaga wyjaśnić, dlaczego jego RNA wydaje się obfite, a białko mniej dominujące. Jednocześnie komórki eksprymujące CHRM1 i CHRM3 miały bardzo podobne sygnatury genowe związane z sygnalizacją synaptyczną i plastycznością, natomiast komórki z CHRM2 tworzyły wyraźnie odrębną grupę powiązaną ze szlakami hamującymi uwalnianie neuroprzekaźników.
Dwa regiony, dwa style dostrajania synaptycznego
Zespół przeszedł następnie od molekuł do funkcji, rejestrując drobne prądy elektryczne z pojedynczych neuronów piramidowych warstwy 3 w preparatach mózgowych. Zanurzyli tkankę w karbacholu — leku aktywującym receptory acetylocholiny — i mierzyli spontaniczne zdarzenia synaptyczne pobudzające i hamujące. W przedniej korze obręczy stymulacja ta zwykle zmniejszała napływ pobudzenia przy jednoczesnym wzroście prądów hamujących, przesuwając lokalną równowagę w stronę silniejszego hamowania aktywności. W bocznej korze przedczołowej natomiast pobudzenie cholinergiczne częściej osłabiało prądy hamujące i pozostawiało relatywnie silniejsze pobudzenie, skłaniając sieć ku stanie bardziej aktywnemu i pobudliwemu. Zmiany te towarzyszyły przekształceniom kształtu i gęstości kolców dendrytycznych — drobnych wypustek gospodarujących synapsami pobudzającymi: karbachol redukował duże, stabilne „grzybkowe” kolce i zwiększał liczbę cieńszych, bardziej plastycznych, przy nieco odmiennych przebiegach czasowych w obu regionach. 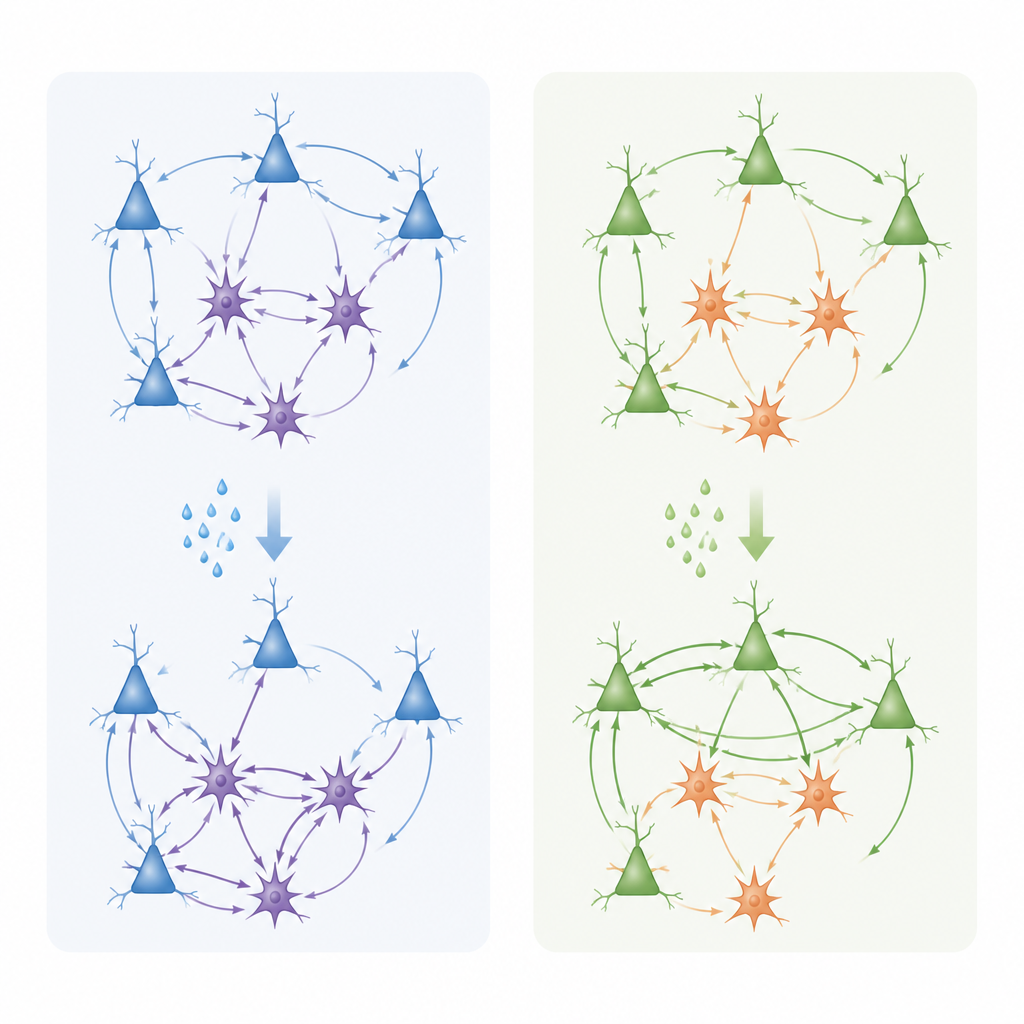
Co to znaczy dla uczenia się i zdrowia psychicznego
Razem wyniki pokazują, że acetylocholina nie działa jak prosty włącznik on–off w korze czołowej. Zamiast tego angażuje różne kombinacje receptorów muskarynowych w określonych typach i warstwach komórek, aby dostroić równowagę między pobudzeniem a hamowaniem w sposób zależny od regionu. W przedniej korze obręczy acetylocholina wydaje się polepszać stosunek sygnału do szumu przez tłumienie napędu pobudzającego i wzmacnianie hamowania, jednocześnie wspierając geny związane z przebudową synaptyczną ważną dla uczenia się emocjonalnego i przetwarzania bólu. W bocznej korze przedczołowej ma skłonność do zachowania lub wzmacniania tonu pobudzającego i promowania strukturalnej elastyczności synaps, co może pomagać w podtrzymaniu pamięci operacyjnej i adaptacyjnego podejmowania decyzji. Łącząc szczegółowe mapy ekspresji genów z rzeczywistymi zmianami aktywności synaptycznej, praca ta dostarcza mechanistycznego ramienia wyjaśniającego, jak zaburzenia cholinergiczne mogą przyczyniać się do objawów poznawczych i afektywnych w zaburzeniach neuropsychiatrycznych, i sugeruje, że ukierunkowanie specyficznych podtypów receptorów muskarynowych w zdefiniowanych obwodach może umożliwić precyzyjniejsze terapie w przyszłości.
Cytowanie: Tsolias, A., Mojica, C.A., Yamani, R. et al. Transcriptional and functional profiles of muscarinic receptor-expressing neurons in primate lateral prefrontal and anterior cingulate cortices. Commun Biol 9, 620 (2026). https://doi.org/10.1038/s42003-026-09866-7
Słowa kluczowe: acetylocholina, kora przedczołowa, przednia kora obręczy, receptory muskarynowe, plastyczność synaptyczna