Clear Sky Science · pl
Wewnątrzwarstwowy mikrokokircuit w skroniowym korze asocjacyjnej leży u podstaw ucieczki wywołanej bodźcem sensorycznym u myszy
Jak mózg przekształca wrażenia w błyskawiczne ucieczki
Gdy mysz gwałtownie ucieka przed głośnym dźwiękiem lub błyskiem światła, jej mózg dokonuje szybkiego, być albo nie być obliczenia: zostać czy uciekać. Badanie to pyta, w którym miejscu mózgu zapada taka decyzja i jak sygnały z oczu, uszu i skóry są łączone w jeden, zdecydowany impuls do ucieczki. Analizując mały obszar kory u myszy, autorzy odsłaniają lokalny schemat połączeń, który wiąże wejścia sensoryczne bezpośrednio z komendą do biegu, dając wskazówki, jak nasze mózgi mogą przekształcać zagrożenie w działanie.
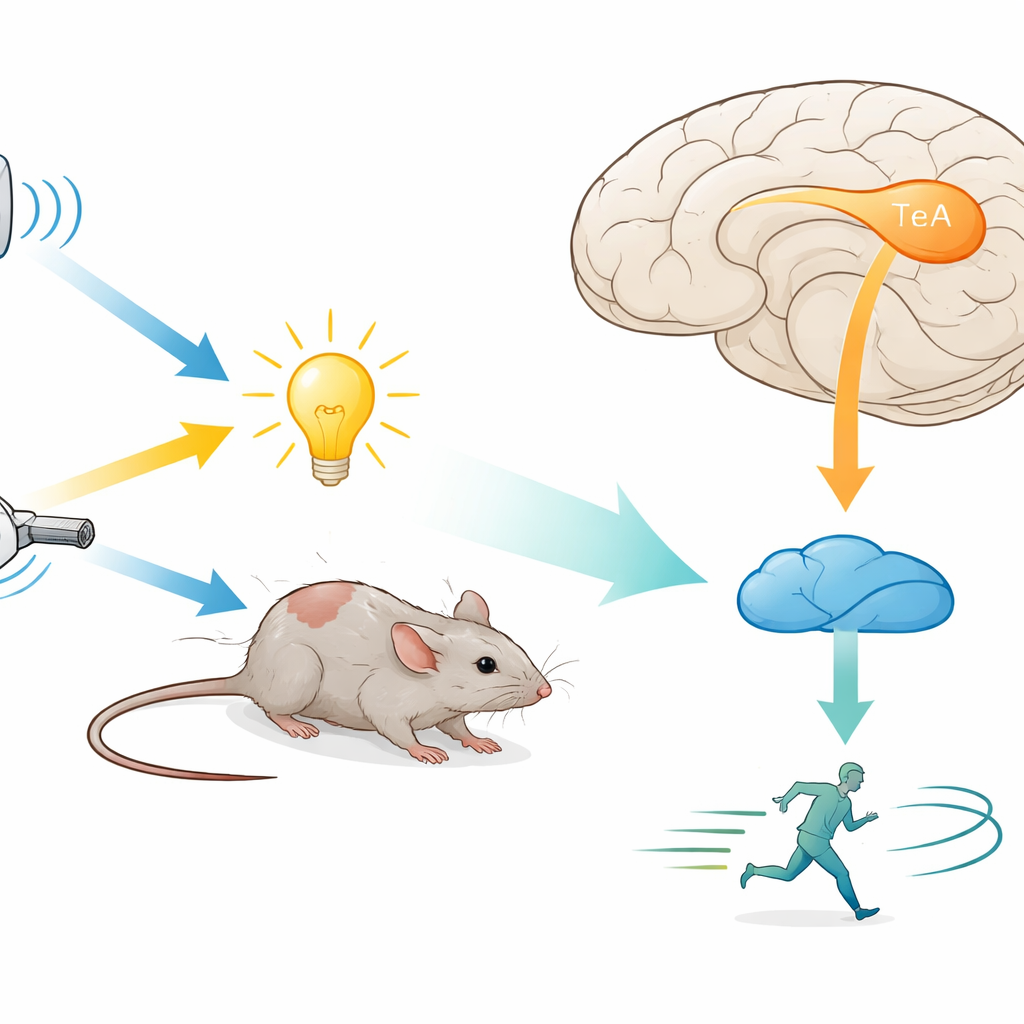
Mały węzeł mózgowy dla wielu rodzajów zagrożeń
Naukowcy skupili się na skroniowej korze asocjacyjnej (TeA), obszarze wyższego rzędu, który otrzymuje informacje z wielu zmysłów i łączy się z regionami kontrolującymi ruch. Umieścili myszy w kontrolowanych warunkach, gdzie można było wywołać nagły dźwięk, światło lub podmuch powietrza. Zarówno w swobodnym pomieszczeniu, jak i na kołowrotku z unieruchomioną głową każdy z tych bodźców niezawodnie wywoływał ucieczkę, przy czym dźwięk był najsilniejszym i najszybszym bodźcem. Gdy zespół tymczasowo uciszał neurony TeA za pomocą projektowanych leków lub hamowania światłem, zachowanie ucieczkowe wobec wszystkich trzech typów bodźców zostało niemal zlikwidowane. To wykazało, że TeA nie jest tylko biernym przekaźnikiem, lecz kluczowym węzłem dla ucieczki, potrzebnym niezależnie od tego, który zmysł pierwszy wykryje zagrożenie.
Z kory do śródmózgowia: bezpośrednia ścieżka ucieczki
Aby ustalić, gdzie TeA wysyła swoje wyjście, autorzy śledzili jego połączenia z użyciem fluorescencyjnych wirusów. Znaleźli gęste projekcje do grzbietowej istoty szarej okołoż wodociągowej (dPAG), regionu śródmózgowia znanego z obecności neuronów „lotu”, które uruchamiają bieg i inne akty obronne. Większość komórek TeA docierających do dPAG była pobudzająca i znajdowała się w cienkim paśmie zwanym warstwą 5a. Wyłączenie tylko tej ścieżki TeA→dPAG, chemicznie lub światłem, blokowało nie tylko ucieczkę wywołaną bodźcem, lecz także zmniejszało normalną spontaniczną aktywność ruchową zwierząt, bez zwiększania lęku. Sugeruje to, że ta droga jest pozytywnym motorem lokomocji — szczególnie w obliczu zagrożenia.
Trzy role neuronów: czucie, decydowanie i wydawanie rozkazu
Wykorzystując drobiazgowe nagrania z pojedynczych komórek TeA u przebudzonych, biegających myszy, zespół wyróżnił trzy funkcjonalne typy neuronów. Jedna grupa reagowała na bodźce wzrokowe, dźwiękowe lub powietrzne, ale wykazywała niewielki związek z prędkością biegu; te neurony działają jako detektory sensoryczne. Druga grupa intensywnie strzelała, gdy zwierzę biegło, ale nie reagowała na pojawienie się bodźców; te komórki kodowały sam rozkaz ruchowy. Trzeci typ robił obie rzeczy: reagował na sygnały sensoryczne, a jego aktywność rosła równocześnie z prędkością biegu myszy. Istotne jest to, że ich wyładowania pojawiały się na kilka sekund przed rozpoczęciem ucieczki, co sugeruje, że pomagają przekształcić „coś się dzieje” w „zacznij biec teraz”.
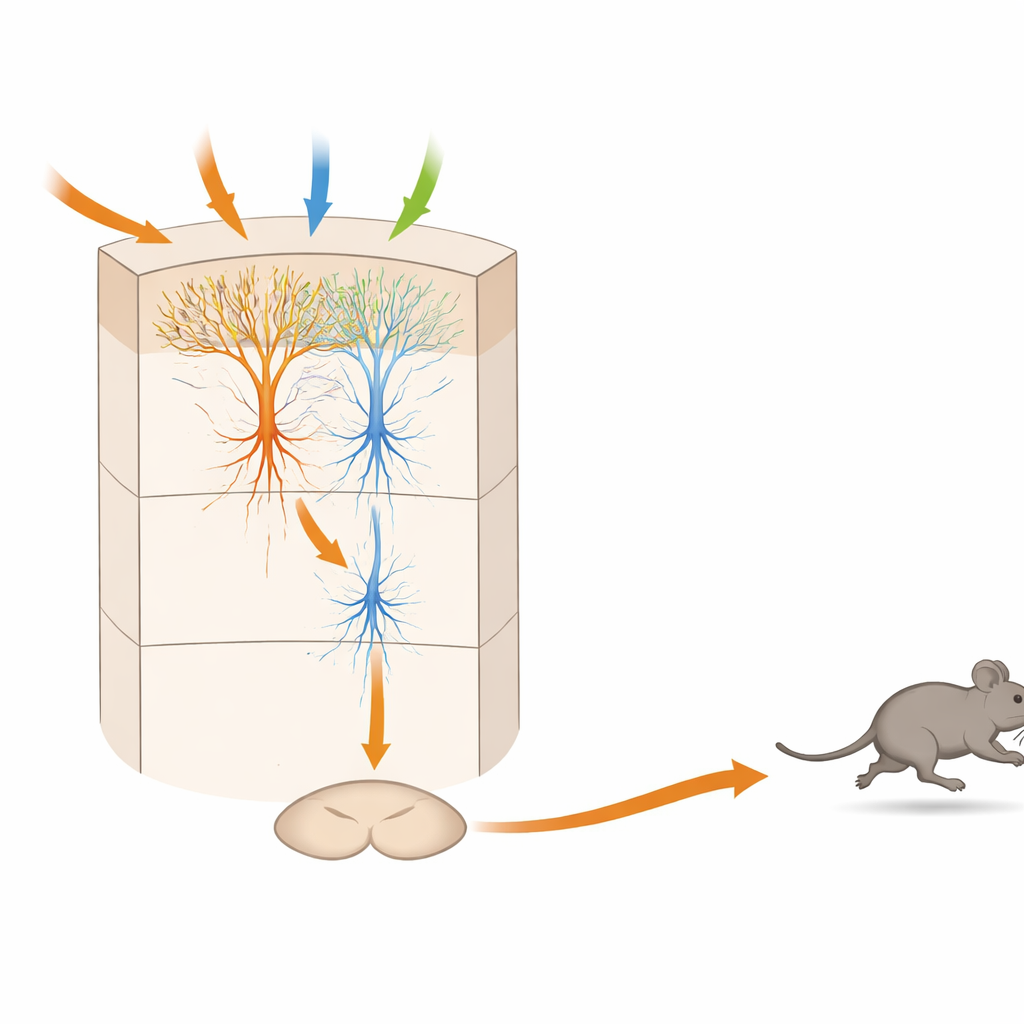
Wielowarstwowy mikrokokircuit, który waży zagrożenie w czasie
Eksperymenty anatomiczne i z preparatami plastra mózgu powiązały te funkcjonalne typy z konkretnymi połączeniami wewnątrz warstwy 5 TeA. Neurony odbierające wejście — „SensTeA”, o grubych rozgałęzionych drzewcach — zbierają sygnały z obszarów związanych ze słuchem, wzrokiem i dotykiem. Wysyłają one bezpośrednie, pobudzające połączenia na bardziej smukłe neurony „TeA_dPAG”, które projektują się do śródmózgowia. Aktywacja komórek po stronie sensorycznej światłem mogła wywołać aktywność w komórkach wyjściowych i przy powtarzanych impulsach ostatecznie wywołać bieg. Połączenie było jednak na tyle słabe, że pojedynczy, krótki wybuch nie wystarczał; zamiast tego aktywność musiała narastać przez setki milisekund do sekund. To temporalne „okno integracji” odpowiada obserwowanemu opóźnieniu między groźnym sygnałem a początkiem ucieczki, sugerując, że obwód akumuluje dowody zanim zaangażuje lot.
Dlaczego to ma znaczenie dla rozumienia decyzji o przeżyciu
Dla laików kluczowy przekaz jest taki, że bardzo mała płatka kory zawiera pełny mini-obwód, który może przyjmować różne ostrzegawcze sygnały sensoryczne, ważyć ich siłę i kombinację, a następnie wydać precyzyjną komendę ruchową do ucieczki. W tym modelu myszy neurony sensoryczne zasilają „neurony decyzyjne”, które z kolei aktywują „neuronypolecenia” połączone bezpośrednio z ośrodkiem ucieczki w śródmózgowiu. Konieczność powtarzającej się aktywności zanim komenda zadziała wyjaśnia, dlaczego istnieje krótkie, ale znaczące opóźnienie między wykryciem zagrożenia a zerwaniem do biegu. Podobna logika może leżeć u podstaw tego, jak ludzki mózg integruje zaszumione, sprzeczne sygnały przed decyzją o ucieczce, zamarciu lub pozostaniu na miejscu, i może informować przyszłe prace nad lękiem, paniką oraz zaburzeniami ruchu, w których ta delikatna równowaga zostaje zaburzona.
Cytowanie: Li, H., Chen, J., Zhong, W. et al. An intralayer microcircuit in the temporal association cortex underlies sensory-induced escape in mice. Nat Commun 17, 4088 (2026). https://doi.org/10.1038/s41467-026-70754-z
Słowa kluczowe: zachowanie ucieczki, integracja sensoryczna, skroniowa kora asocjacyjna, neuronalne mikrokokiruity, lokomocja myszy