Clear Sky Science · es
Un microcircuito intracapa en la corteza de asociación temporal subyace a la huida inducida por estímulos sensoriales en ratones
Cómo el cerebro convierte los sentidos en escapadas en fracciones de segundo
Cuando un ratón sale disparado ante un ruido fuerte o un destello de luz, su cerebro está realizando un cálculo rápido de vida o muerte: quedarse o correr. Este estudio pregunta dónde en el cerebro se toma esa decisión y cómo las señales de los ojos, oídos y piel se integran en una única orden decisiva de huida. Al diseccionar una pequeña región de la corteza en ratones, los autores descubren un diagrama de conexiones local que enlaza la entrada sensorial directamente con la orden de correr, ofreciendo pistas de cómo nuestros propios cerebros podrían convertir el peligro en acción.
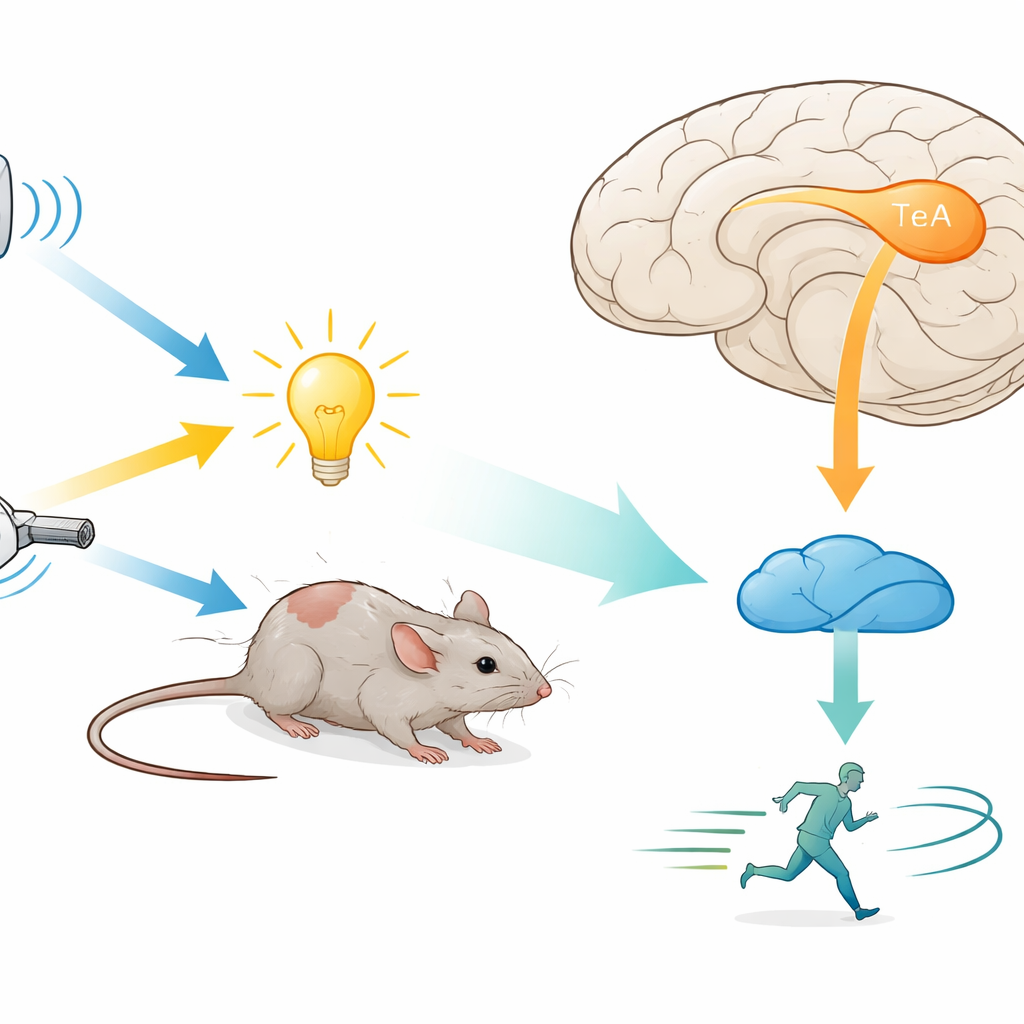
Un pequeño centro cerebral para muchos tipos de amenazas
Los investigadores se centraron en la corteza de asociación temporal (TeA), una área de orden superior que recibe información de varios sentidos y se conecta con regiones que controlan el movimiento. Colocaron a los ratones en entornos controlados donde podían administrarse de forma repentina sonidos, luces o soplos de aire. Tanto en una arena con libre movimiento como en una rueda de correr con la cabeza fijada, cada uno de estos estímulos hacía que los animales huyeran de manera fiable, siendo el sonido el desencadenante más potente y más rápido. Cuando el equipo silenció temporalmente las neuronas de la TeA usando fármacos diseñados o inhibición dependiente de luz, el comportamiento de huida ante los tres tipos de estímulo quedó casi abolido. Esto mostró que la TeA no es solo un relevo pasivo, sino un centro crucial para la huida, necesario independientemente de qué sentido detecte primero la amenaza.
De la corteza al mesencéfalo: una vía directa de escape
Para ver a dónde envía su salida la TeA, los autores trazaron sus conexiones usando virus fluorescentes. Encontraron una proyección densa hacia la sustancia gris periacueductal dorsal (dPAG), una región mesencefálica conocida desde hace tiempo por albergar neuronas de “huida” que impulsan la carrera y otros actos defensivos. La mayoría de las células de la TeA que llegaban a la dPAG eran excitadoras y se ubicaban en una banda delgada llamada capa 5a. Desactivar únicamente esta vía TeA→dPAG, ya fuera químicamente o con luz, bloqueó no solo la huida evoked por estímulos sino que también redujo el movimiento espontáneo normal de los animales, sin aumentar la ansiedad. Esto sugiere que la vía es un impulsor positivo de la locomoción, especialmente cuando hay peligro.
Tres roles neuronales: detectar, decidir y mandar
Mediante registros de alta resolución de células individuales de la TeA en ratones despiertos y en movimiento, el equipo identificó tres tipos funcionales de neuronas. Un grupo respondía a vistas, sonidos o soplos de aire pero mostraba poca relación con la velocidad de carrera; estas neuronas actúan como detectores sensoriales. Un segundo grupo disparaba fuertemente cuando el animal corría pero no cuando aparecían estímulos; estas células codificaban la orden motora en sí. El tercer grupo hacía ambas cosas: respondía a las señales sensoriales y su actividad aumentaba en paralelo con la velocidad a la que corría el ratón. Es importante que sus picos de actividad tendían a ocurrir un par de segundos antes del inicio de la huida, lo que implica que ayudan a transformar “algo está ocurriendo” en “empieza a correr ahora”.
Un microcircuito estratificado que pondera el peligro a lo largo del tiempo
Experimentos anatómicos y de fisiología en cortes cerebrales relacionaron luego estos tipos funcionales con cableados específicos dentro de la capa 5 de la TeA. Las neuronas receptoras de entrada, “SensTeA”, que son de gran tronco y muy ramificadas, recogen señales de áreas auditivas, visuales y táctiles. Envían conexiones excitadoras directas hacia neuronas más esbeltas “TeAdPAG” que proyectan hacia el mesencéfalo. Activar con luz las células del lado sensorial podía impulsar la actividad en las células de salida y, con pulsos repetidos, eventualmente provocar la carrera. Sin embargo, la conexión era lo bastante débil como para que un único estallido breve no fuera suficiente; en su lugar, la actividad debía acumularse durante cientos de milisegundos a segundos. Esta “ventana de integración” temporal coincide con el retraso observado entre una señal amenazante y el inicio de la huida, lo que sugiere que el circuito acumula evidencia antes de comprometerse a la fuga.
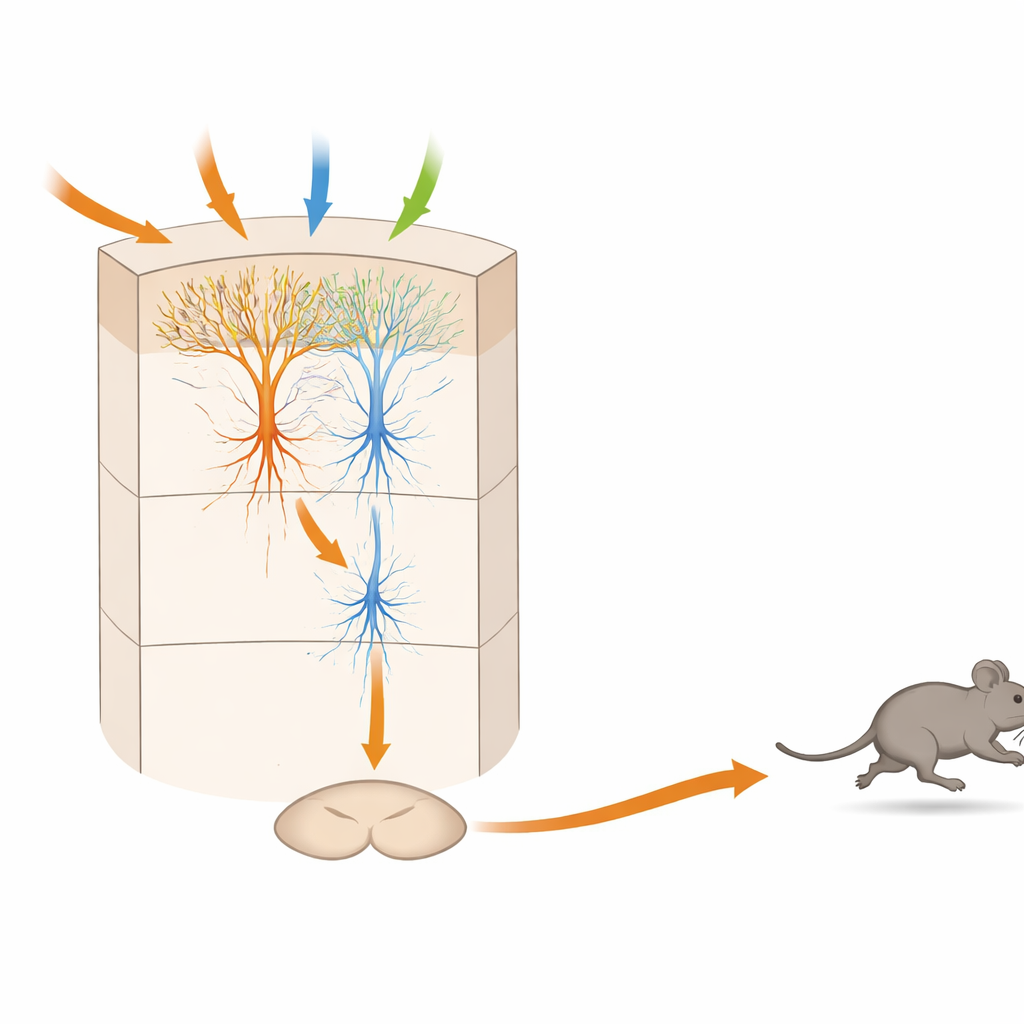
Por qué esto importa para entender las decisiones de supervivencia
Para un público no especializado, el mensaje clave es que un parche muy pequeño de la corteza contiene un minicircuito completo que puede recibir distintas advertencias sensoriales, ponderar su intensidad y combinación, y emitir luego una orden motora precisa para correr. En este modelo de ratón, las neuronas sensoriales alimentan a neuronas de “decisión”, que a su vez activan neuronas de “comando” conectadas directamente a un centro de escape mesencefálico. La necesidad de actividad repetida antes de que se dispare la orden explica por qué existe un breve pero significativo retraso entre detectar el peligro y echar a correr. Una lógica similar podría subyacer en cómo los cerebros humanos integran señales ruidosas o en conflicto antes de decidir huir, quedarse paralizados o permanecer en el sitio, y podría orientar trabajos futuros sobre ansiedad, pánico y trastornos del movimiento donde este delicado equilibrio falla.
Cita: Li, H., Chen, J., Zhong, W. et al. An intralayer microcircuit in the temporal association cortex underlies sensory-induced escape in mice. Nat Commun 17, 4088 (2026). https://doi.org/10.1038/s41467-026-70754-z
Palabras clave: comportamiento de huida, integración sensorial, corteza de asociación temporal, microcircuitos neuronales, locomoción en ratones