Clear Sky Science · it
Un microcircuito intrastrato nella corteccia temporale associativa sottende la fuga indotta da stimoli sensoriali nei topi
Come il cervello trasforma i sensi in fughe in una frazione di secondo
Quando un topo scatta via all'improvviso per un rumore forte o un lampo di luce, il suo cervello compie un rapido calcolo di vita o di morte: restare o fuggire. Questo studio indaga dove nel cervello viene presa quella decisione e come i segnali provenienti da occhi, orecchie e pelle vengono trasformati in un comando unitario e risoluto di fuga. Dissecando una piccola regione della corteccia nei topi, gli autori scoprono uno schema di connessioni locali che collega direttamente l'input sensoriale al comando di corsa, offrendo indizi su come anche i nostri cervelli potrebbero tradurre il pericolo in azione.
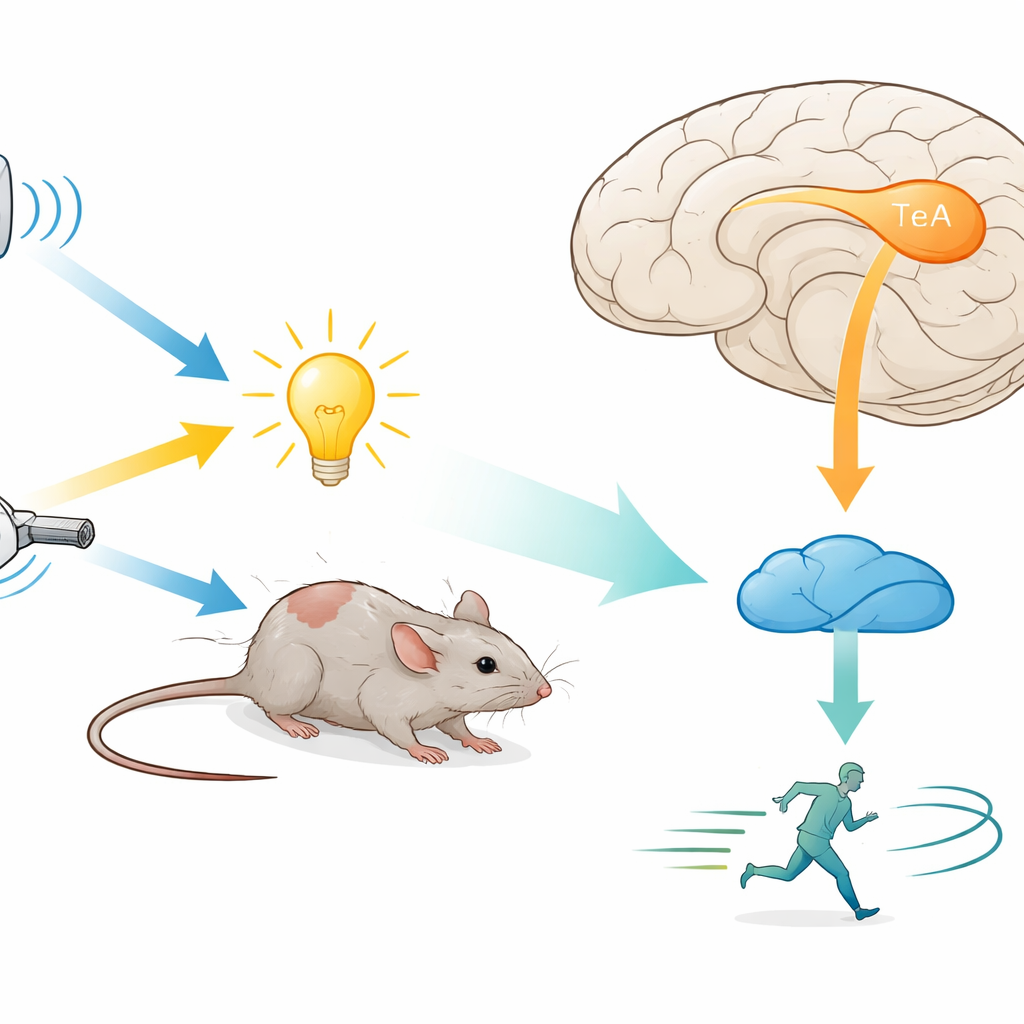
Un piccolo centro cerebrale per molti tipi di minaccia
I ricercatori si sono concentrati sulla corteccia temporale associativa (TeA), un'area di ordine superiore che riceve informazioni da più sensi e si collega a regioni che controllano il movimento. Hanno posto i topi in ambienti controllati dove potevano essere somministrati all'improvviso suoni, luci o getti d'aria. Sia nell'arena a movimento libero sia sulla ruota per animali con la testa fissata, ciascuno di questi segnali induceva costantemente la fuga, con il suono come stimolo più potente e rapido. Quando il team ha temporaneamente silenziato i neuroni della TeA usando droghe designer o inibizione controllata dalla luce, il comportamento di fuga verso tutti e tre i tipi di stimolo è quasi scomparso. Ciò mostra che la TeA non è solo un relè passivo ma un nodo cruciale per la fuga, necessario indipendentemente da quale senso per primo rilevi la minaccia.
Dalla corteccia al mesencefalo: una via diretta per la fuga
Per vedere dove la TeA invia i suoi output, gli autori hanno tracciato le sue connessioni usando virus fluorescenti. Hanno trovato una densa proiezione verso la sostanza grigia periacqueduttale dorsale (dPAG), una regione del mesencefalo nota da tempo per ospitare neuroni di “fuga” che promuovono la corsa e altri atti difensivi. La maggior parte delle cellule della TeA che raggiungevano la dPAG era eccitatoria e si trovava in una sottile banda chiamata strato 5a. Spegnere solo questo percorso TeA→dPAG, chimicamente o con la luce, bloccava non solo la fuga evocata dallo stimolo ma riduceva anche il movimento spontaneo normale degli animali, senza aumentare l'ansia. Ciò suggerisce che questa via è un motore positivo della locomozione, specialmente in presenza di pericolo.
Tre ruoli neuronali: percepire, decidere e comandare
Usando registrazioni ad alta risoluzione da singole cellule della TeA in topi svegli e in corsa, il team ha identificato tre tipi funzionali di neuroni. Un gruppo rispondeva a stimoli visivi, sonori o a getti d'aria ma mostrava poca correlazione con la velocità di corsa; questi neuroni fungono da rivelatori sensoriali. Un secondo gruppo si attivava in modo marcato quando l'animale correva ma non alla comparsa degli stimoli; queste cellule codificano il comando motorio stesso. Il terzo gruppo faceva entrambe le cose: rispondeva agli indizi sensoriali e la sua attività aumentava in parallelo con la velocità di corsa. È importante che i loro spike tendevano a verificarsi qualche decina di millisecondi fino a un paio di secondi prima dell'inizio della fuga, il che suggerisce che aiutano a trasformare «sta succedendo qualcosa» in «inizia a correre adesso».
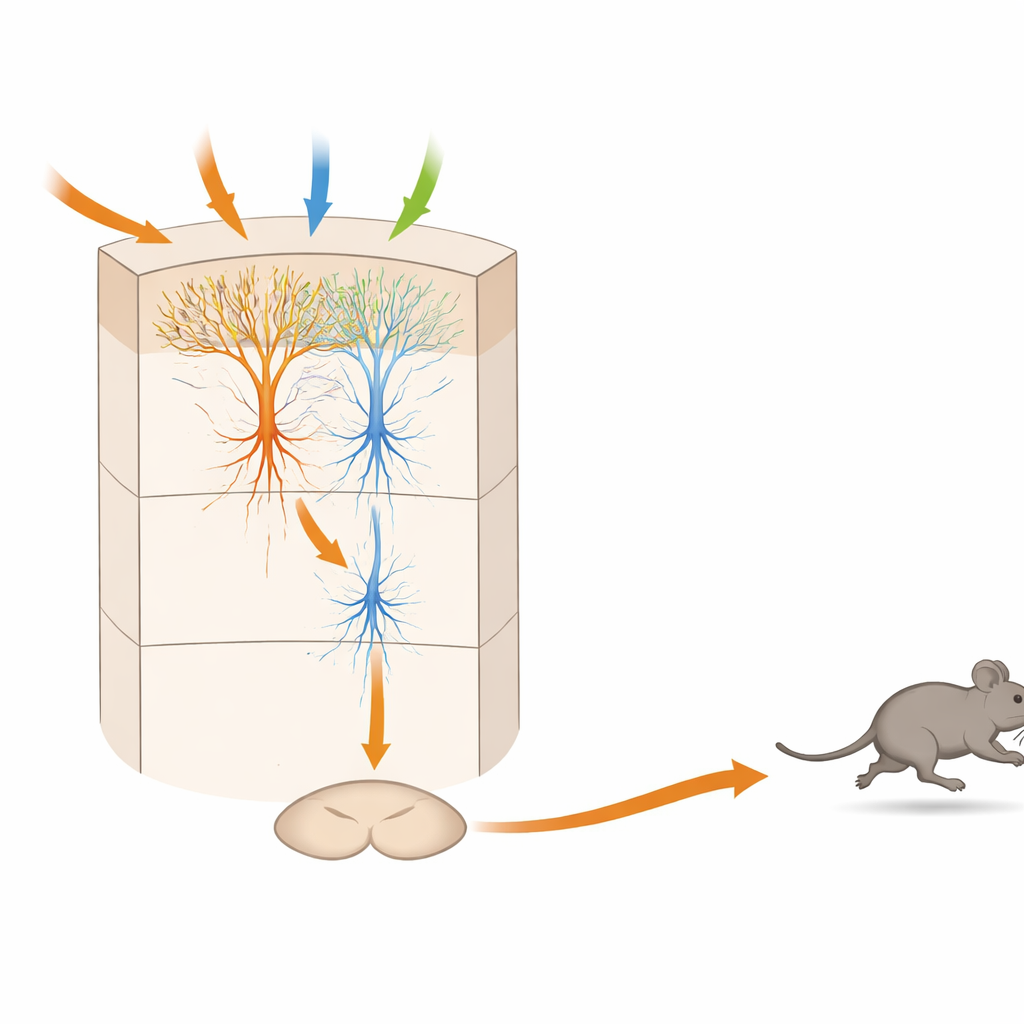
Un microcircuito stratificato che pesa il pericolo nel tempo
Esperimenti anatomici e di fisiologia su fette cerebrali hanno poi collegato questi tipi funzionali a connessioni specifiche all'interno dello strato 5 della TeA. I neuroni riceventi di input, detti “SensTeA”, con grandi dendriti a ciuffo e ampia ramificazione, raccolgono segnali dalle aree uditive, visive e tattili. Essi inviano connessioni eccitatorie dirette a neuroni più snelli chiamati “TeAdPAG” che proiettano verso il mesencefalo. Attivare con la luce le cellule del lato sensoriale poteva indurre attività nelle cellule di output e, con impulsi ripetuti, alla fine scatenare la corsa. Tuttavia, la connessione era sufficientemente debole perché una singola scarica breve non bastasse; invece, l'attività doveva accumularsi per centinaia di millisecondi fino a secondi. Questa «finestra di integrazione» temporale corrisponde al ritardo osservato tra un segnale minaccioso e l'inizio della fuga, suggerendo che il circuito accumula prove prima di impegnarsi nella fuga.
Perché questo è importante per comprendere le decisioni di sopravvivenza
Per un non specialista, il messaggio chiave è che una piccolissima porzione di corteccia contiene un mini‑circuito completo in grado di ricevere diversi avvisi sensoriali, valutarne l'intensità e la combinazione, e poi emettere un comando motorio preciso per correre. In questo modello murino, i neuroni sensoriali alimentano i neuroni di «decisione», che a loro volta attivano i neuroni di «comando» connessi direttamente a un centro di fuga nel mesencefalo. La necessità di un'attività ripetuta prima che il comando si accenda spiega perché esiste un breve ma significativo ritardo tra la percezione del pericolo e la fuga. Una logica analoga potrebbe sottendere il modo in cui il cervello umano integra segnali rumorosi o contrastanti prima di decidere se fuggire, immobilizzarsi o restare, e potrebbe orientare lavori futuri su ansia, panico e disturbi del movimento in cui questo equilibrio delicato va in tilt.
Citazione: Li, H., Chen, J., Zhong, W. et al. An intralayer microcircuit in the temporal association cortex underlies sensory-induced escape in mice. Nat Commun 17, 4088 (2026). https://doi.org/10.1038/s41467-026-70754-z
Parole chiave: comportamento di fuga, integrazione sensoriale, corteccia temporale associativa, microcircuiti neurali, locomozione nei topi