Clear Sky Science · pl
Globalny sygnał błędu kieruje lokalną optymalizacją w obliczaniu niedopasowania
Dlaczego mózg nie płoszy się własnymi działaniami
Gdy klikniesz myszką, żeby odtworzyć piosenkę, nie dziwi cię dźwięk, który następuje. Jeśli jednak muzyka zaczyna się niespodziewanie bez twojego dotyku, przykuwa twoją uwagę. Ta codzienna różnica między oczekiwanymi a nieoczekiwanymi wrażeniami zależy od zdolności mózgu do przewidywania sensorycznych następstw naszych ruchów i wyeksponowania tylko niezgodności. Artykuł bada, jak sieci neuronów mogą się nauczyć dokonywać takich porównań w sposób niezawodny i co idzie nie tak, gdy to uczenie zostaje zaburzone.
Jak mózg przewiduje własny świat sensoryczny
Naukowcy od dawna wiedzą, że mózg wysyła wewnętrzną kopię poleceń ruchowych, zwaną kopiowaniem korolarycznym, do obszarów sensorycznych. Sygnał ten niesie przewidywanie, co powinniśmy zobaczyć lub usłyszeć podczas ruchu. Jeśli napływający bodziec ze świata zewnętrznego zgadza się z tą prognozą, odpowiedzi sensoryczne są tłumione; jeśli nie, pewne neurony odpowiadają silnie, sygnalizując błąd predykcji. Autorzy koncentrują się na dwóch rodzajach takich neuronów: tych, które reagują, gdy bodziec jest silniejszy niż przewidywano (pozytywny błąd predykcji), oraz tych, które reagują, gdy przewidywanie przewyższa rzeczywisty bodziec (negatywny błąd predykcji). Razem pozwalają mózgowi wykrywać zarówno brakujące, jak i niespodziewane wrażenia, na przykład dźwięk, który nie następuje przy naszym działaniu, albo dźwięk pojawiający się bez odpowiadającego ruchu. 
Globalny sygnał uczący dla lokalnego okablowania
Pierwsze spojrzenie sugeruje, że obliczenie niedopasowania jest proste: odjąć przewidywanie od bodźca i wziąć wartość bezwzględną. Jednak rzeczywiste obwody korowe napotykają kilka przeszkód. Połączenia długodystansowe między obszarami mózgu są głównie pobudzające, podczas gdy do znoszenia tych wejść potrzebne są lokalne komórki hamujące. Ponadto neurony w korze sensorycznej mają tendencję do niskich stężeń wyjściowych w stanie podstawowym, co utrudnia reprezentowanie zarówno dodatnich, jak i ujemnych odchyleń. Centralna propozycja tej pracy to biologicznie wiarygodna reguła uczenia „trzyczynnikowego”, która pozwala lokalnym synapsom hamującym przebudowywać się z użyciem pojedynczego globalnego sygnału uczącego. Sygnał ten, przypuszczalnie przenoszony przez neuromodulatory takie jak noradrenalina z pnia mózgu, odzwierciedla ogólną wielkość niedopasowania, bez konieczności rozpoznawania, która konkretna synapsa je spowodowała.
Jak lokalne obwody uczą się sygnalizować zaskoczenie
W modelu każdy neuron piramidalny otrzymuje zarówno wejścia sensoryczne, jak i przewidywania, a także zahamowanie przekazywane przez lokalne interneurony. Początkowo zahamowanie jest w przybliżeniu jednorodne i nie usuwa właściwych wejść, więc oczekiwane zdarzenia nadal wywołują silne odpowiedzi. W trakcie uczenia, gdy przewidywanie i bodziec zazwyczaj się zgadzają, globalny sygnał ma jeden znak i napędza korektę podobną do reguły Hebba, która wyrównuje całkowite pobudzenie i zahamowanie, przesuwając system na „linię wolnego uczenia”, gdzie ogólna aktywność jest niska w oczekiwanych warunkach. W rzadziej występujących próbach z niedopasowaniem znak globalnego sygnału odwraca się, a ta sama lokalna reguła staje się anty-Hebbiańska, popychając wagi synaptyczne wzdłuż tej linii, aż każdy neuron stanie się wyspecjalizowany. Jedna podgrupa staje się neuronami pozytywnego błędu predykcji, pobudzanymi głównie gdy bodziec przewyższa przewidywanie; druga staje się neuronami negatywnego błędu predykcji, reagującymi, gdy przewidywanie przewyższa bodziec. Autorzy pokazują matematycznie, że ta prosta reguła zbiega do tego samego optymalnego rozwiązania co metoda spadku gradientu, ale używając wyłącznie biologicznie realistycznych składników. 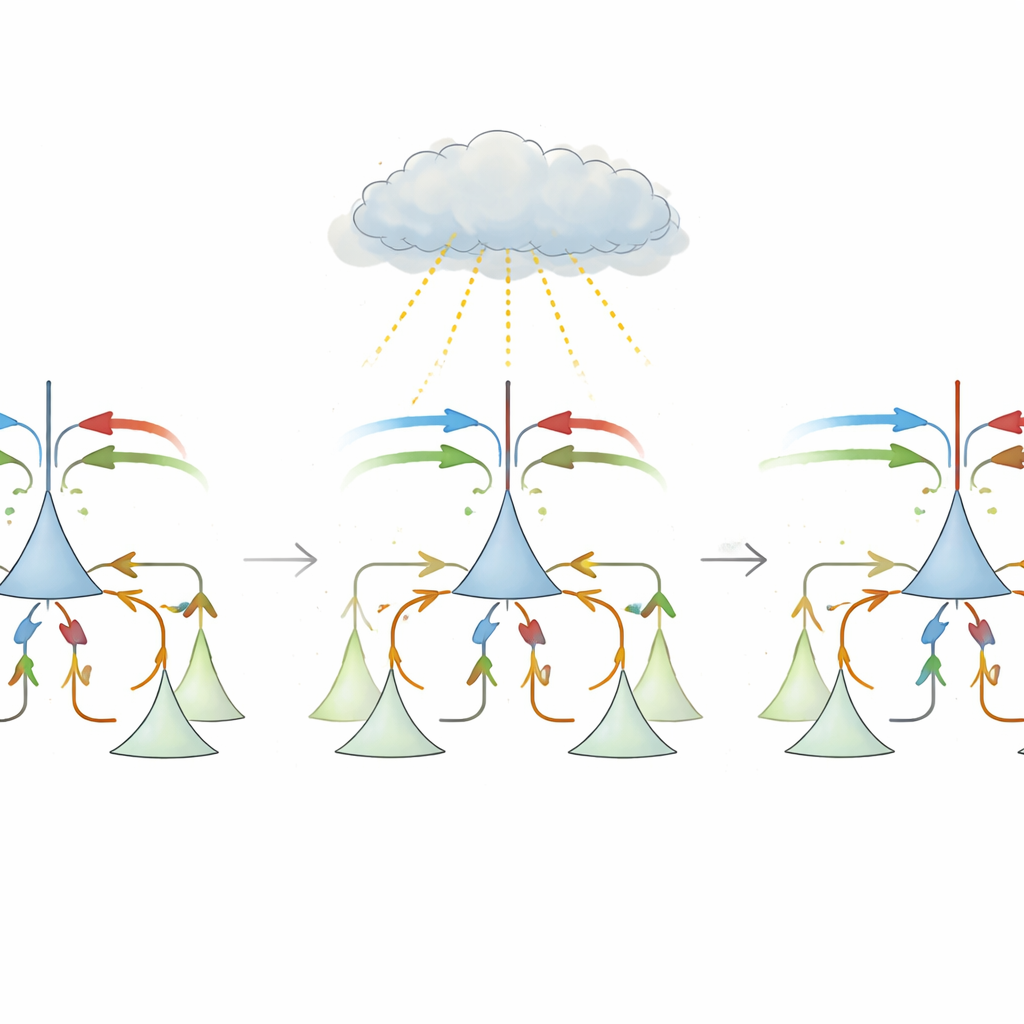
Powiązanie modelu z danymi z prawdziwego mózgu
Następnie badacze przechodzą od uproszczonych jednostek do bardziej realistycznych modeli neuronów i testują, czy wyuczone obwody odtwarzają znane eksperymenty na myszach. W jednym paradygmacie myszy uczą się, że bieganie na bieżni wywołuje ruch wizualny; później krótkie zatrzymanie sceny wizualnej podczas ruchu powoduje silną odpowiedź niedopasowania w korze wzrokowej. Model pokazuje ten sam wzorzec: po nauce aktywność przy oczekiwanym ruchu jest stłumiona, podczas gdy nagłe pauzy wywołują duże odpowiedzi, szczególnie w neuronach negatywnego błędu predykcji, które normalnie są hamowane przez wejście wzrokowe. Gdy autorzy symulują zakłócenia różnych klas komórek hamujących lub blokadę receptorów NMDA wspierających plastyczność, zdolność sieci do sygnalizowania niedopasowania pogarsza się w sposób zgodny z wynikami farmakologicznymi i optogenetycznymi. Na koniec model przewiduje, że po nauce neurony powinny podzielić się na dwie grupy o przeciwnych korelacjach z siłą bodźca i siłą przewidywania. Ponowna analiza istniejących danych mysich potwierdza to: tylko zwierzęta z właściwym sprzężeniem ruch–wzrok wykazują wyraźny wzorzec bimodalny, zgodny z odrębnymi populacjami pozytywnych i negatywnych neuronów błędu predykcji.
Dlaczego to ma znaczenie dla percepcji i zdrowia psychicznego
Badanie wspiera ujednoliconą koncepcję, w której globalne, rozsyłane po mózgu sygnały błędu dostrajają bardzo konkretne lokalne połączenia, dzięki czemu obszary sensoryczne stają się ekspertami w tłumieniu przewidywalnego i uwydatnianiu zaskakującego. W codziennym życiu sprawia to, że nasza percepcja jest stabilna podczas własnych ruchów, a jednocześnie pozostajemy wrażliwi na nieoczekiwane zdarzenia. Ta sama mechanika może także odciążyć wyższe obszary mózgu z pracy nad szczegółowymi przewidywaniami, uwalniając zasoby poznawcze po opanowaniu umiejętności. Ponieważ zaburzenia kopiowania korolarycznego i sygnalizacji błędu predykcji łączone są z halucynacjami i urojeniami, proponowany mechanizm oferuje też konkretne ramy obwodów neuronalnych do zrozumienia, jak takie objawy mogą się pojawić, gdy uczenie hamujące lub neuromodulacyjne sygnały błędu zawodzą.
Cytowanie: Meng, J.H., Wang, XJ. Global error signal guides local optimization in mismatch calculation. Nat Commun 17, 3868 (2026). https://doi.org/10.1038/s41467-026-70354-x
Słowa kluczowe: kodowanie predykcyjne, kopiowanie korolaryczne, plastyczność hamująca, neuromodulacja, neurony sygnalizujące błąd predykcji