Clear Sky Science · es
Señal de error global guía la optimización local en el cálculo de desajustes
Por qué nuestro cerebro no se sobresalta con nuestras propias acciones
Cuando haces clic en un ratón para reproducir una canción, no te sorprende el sonido que sigue. Pero si la música comienza de repente sin que la hayas tocado, capta tu atención. Esta diferencia cotidiana entre sensaciones esperadas e inesperadas se basa en la capacidad del cerebro para predecir las consecuencias sensoriales de nuestros propios movimientos y para resaltar solo las discrepancias. Este artículo explora cómo las redes neuronales podrían aprender a hacer esas comparaciones de forma fiable y qué falla cuando ese aprendizaje se ve interrumpido.
Cómo el cerebro predice su propio mundo sensorial
Los científicos saben desde hace tiempo que el cerebro envía una copia interna de las órdenes de movimiento, llamada descarga corolaria, a las áreas sensoriales. Esta señal transporta una predicción de lo que deberíamos ver u oír cuando nos movemos. Si el estímulo entrante del mundo exterior coincide con esta predicción, las respuestas sensoriales se atenuan; si no, ciertas neuronas responden con fuerza, señalando un error de predicción. Los autores se centran en dos tipos de dichas neuronas: las que responden cuando el estímulo es más fuerte de lo predicho (error de predicción positivo) y las que responden cuando la predicción es mayor que el estímulo real (error de predicción negativo). En conjunto, permiten al cerebro detectar tanto las sensaciones ausentes como las inesperadas, como un sonido que no ocurre cuando actuamos o un sonido que aparece sin un movimiento coincidente. 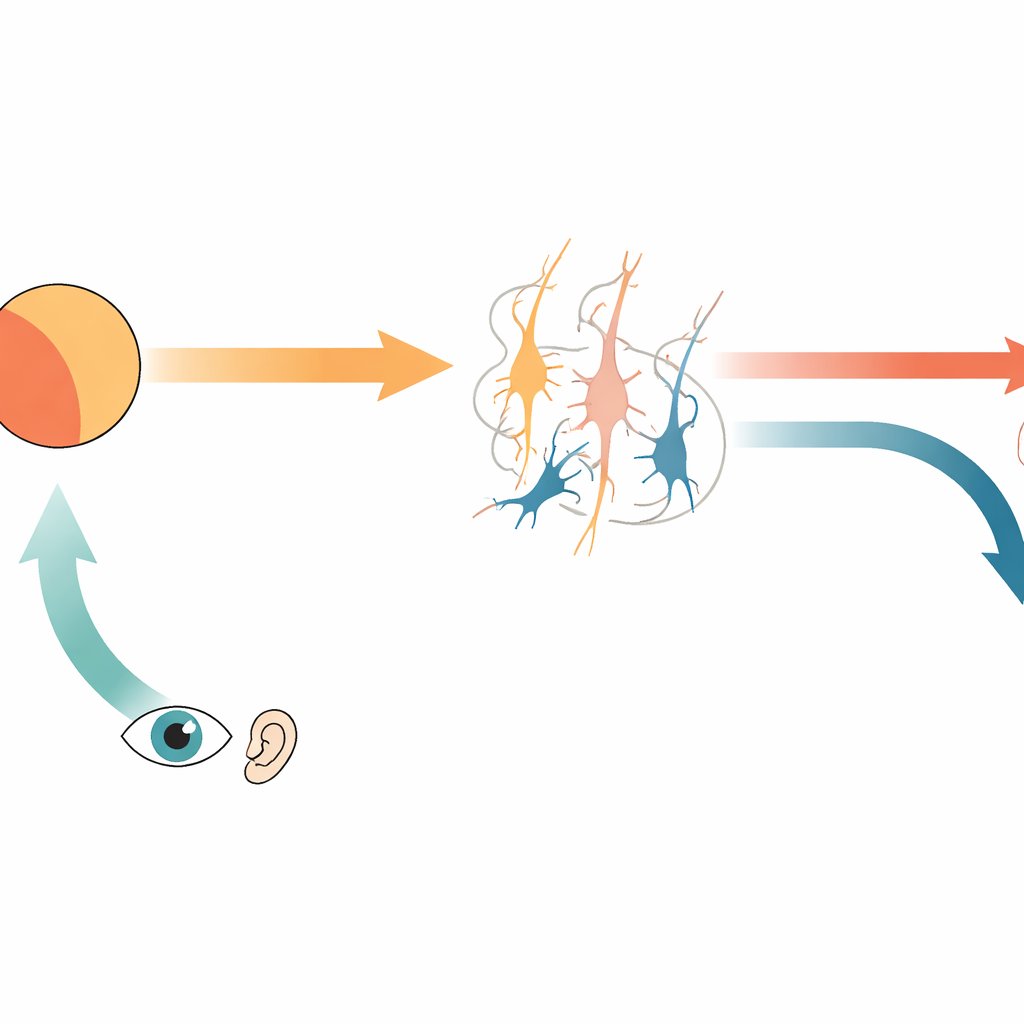
Una señal global de enseñanza para el cableado local
A primera vista, calcular un desajuste parece sencillo: restar la predicción al estímulo y tomar el valor absoluto. Pero los circuitos corticales reales enfrentan varios obstáculos. Las conexiones de largo alcance entre áreas cerebrales son mayoritariamente excitatorias, mientras que las células inhibitorias locales son necesarias para cancelar entradas. Además, las neuronas en la corteza sensorial tienden a disparar a tasas de referencia bajas, lo que complica representar tanto las desviaciones positivas como las negativas. La propuesta central de este trabajo es una regla de aprendizaje “de tres factores” biológicamente plausible que permite a las sinapsis inhibitorias locales reconfigurarse usando una única señal global de enseñanza. Esta señal, que se piensa que es transportada por neuromoduladores como la noradrenalina desde el tronco encefálico, refleja cuán grande es el desajuste en conjunto, sin necesidad de saber qué sinapsis específica lo causó.
Cómo los circuitos locales aprenden a señalar la sorpresa
En el modelo, cada neurona piramidal recibe tanto entradas de estímulo como de predicción, además de inhibición canalizada a través de interneuronas locales. Inicialmente, la inhibición es aproximadamente uniforme y no logra cancelar las entradas correctas, por lo que los eventos esperados siguen evocando respuestas fuertes. Durante el aprendizaje, cuando predicción y estímulo suelen coincidir, la señal global tiene un signo y provoca un ajuste tipo Hebbiano que equilibra la excitación y la inhibición totales, empujando el sistema hacia una “línea de aprendizaje lento” donde la actividad global es baja en condiciones esperadas. En pruebas de desajuste más raras, el signo de la señal global se invierte y la misma regla local se vuelve anti-Hebbiana, empujando los pesos sinápticos a lo largo de esa línea hasta que cada neurona termina especializada. Un subgrupo se convierte en neuronas de error de predicción positivo, excitadas principalmente cuando el estímulo supera a la predicción; otro se convierte en neuronas de error de predicción negativo, que responden cuando la predicción sobrepasa al estímulo. Los autores muestran matemáticamente que esta regla simple converge a la misma solución óptima que el descenso de gradiente, pero usando solo ingredientes biológicamente realistas. 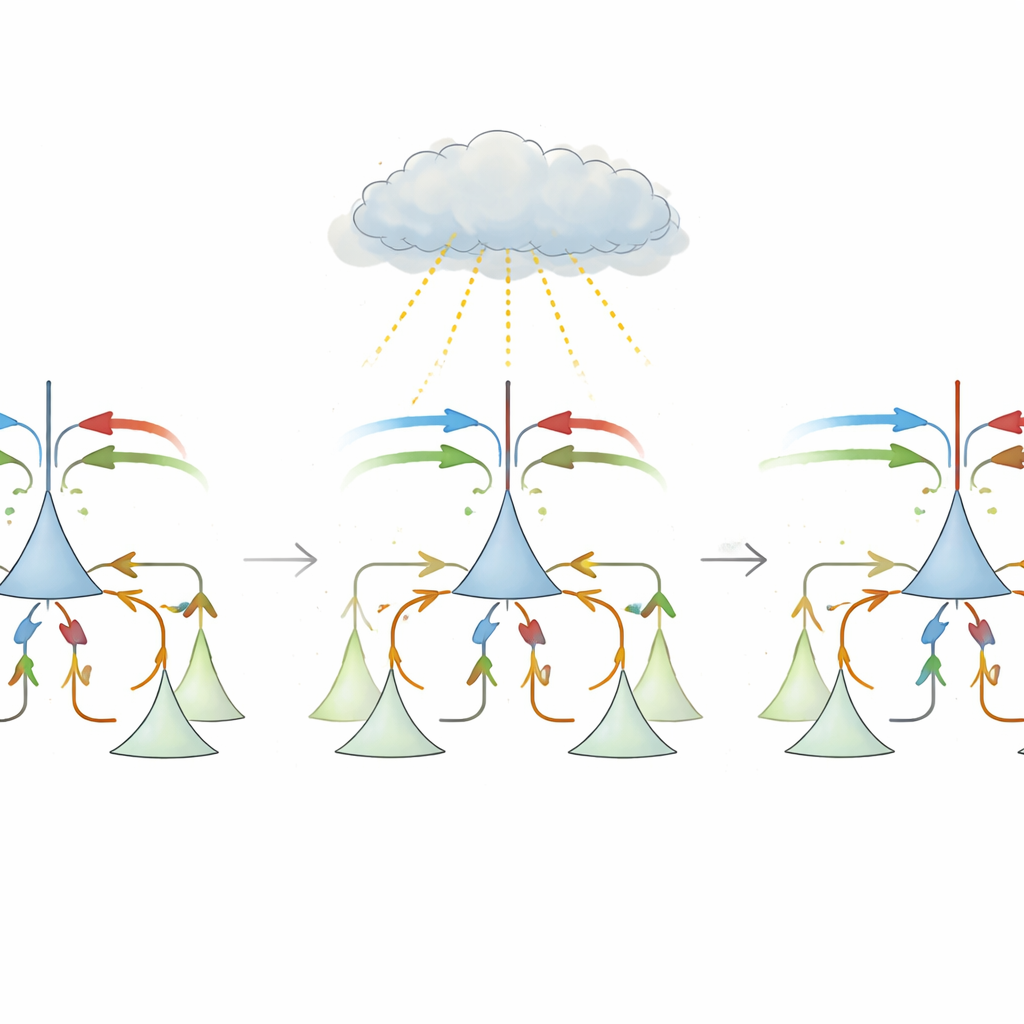
Vinculando el modelo con datos reales del cerebro
Los investigadores pasan luego de unidades simplificadas a modelos neuronales más realistas y ponen a prueba si los circuitos aprendidos reproducen experimentos conocidos en ratones. En un paradigma, los ratones aprenden que correr en una cinta produce movimiento visual; más tarde, detener brevemente la escena visual durante el movimiento provoca una fuerte respuesta de desajuste en la corteza visual. El modelo muestra el mismo patrón: tras el aprendizaje, la actividad ante el movimiento esperado queda suprimida, mientras que las pausas repentinas evocan grandes respuestas, especialmente en las neuronas de error de predicción negativo que normalmente quedan frenadas por la entrada visual. Cuando los autores simulan la perturbación de distintas clases de células inhibitorias, o el bloqueo de los receptores NMDA que sostienen la plasticidad, la capacidad de la red para señalar desajustes se degrada de maneras que concuerdan con hallazgos farmacológicos y optogenéticos. Finalmente, el modelo predice que, tras el aprendizaje, las neuronas deberían dividirse en dos grupos con correlaciones opuestas respecto a la fuerza del estímulo y de la predicción. Un reanálisis de datos existentes en ratones confirma esto: solo los animales con un acoplamiento correcto entre movimiento y visión muestran un patrón claramente bimodal, consistente con poblaciones distintas de errores de predicción positivos y negativos.
Por qué esto importa para la percepción y la salud mental
El estudio respalda una visión unificadora en la que una difusión global de error en todo el cerebro afina conexiones locales muy específicas para que las áreas sensoriales se vuelvan expertas en cancelar lo predecible y destacar lo sorprendente. En la vida cotidiana, esto hace que nuestra percepción sea estable durante nuestros propios movimientos al tiempo que nos mantiene sensibles a eventos inesperados. La misma maquinaria también puede ayudar a aliviar a regiones cerebrales superiores del trabajo de predicción detallada, liberando recursos cognitivos una vez que se han aprendido las habilidades. Dado que las alteraciones de la descarga corolaria y de la señalización del error de predicción se han vinculado con alucinaciones y delirios, el mecanismo propuesto ofrece además un marco de circuito neural concreto para entender cómo podrían surgir tales síntomas cuando el aprendizaje inhibitorio o las señales neuromoduladoras de error fallan.
Cita: Meng, J.H., Wang, XJ. Global error signal guides local optimization in mismatch calculation. Nat Commun 17, 3868 (2026). https://doi.org/10.1038/s41467-026-70354-x
Palabras clave: codificación predictiva, descarga corolaria, plasticidad inhibitoria, neuromodulación, neuronas de error de predicción