Clear Sky Science · fr
Le signal d’erreur global guide l’optimisation locale dans le calcul des écarts
Pourquoi notre cerveau ne sursaute pas à nos propres actions
Quand vous cliquez sur une souris pour lancer une chanson, le son qui suit ne vous surprend pas. Mais si la musique démarre soudainement sans votre geste, elle capte votre attention. Cette différence quotidienne entre sensations attendues et inattendues repose sur la capacité du cerveau à prédire les conséquences sensorielles de nos mouvements et à ne mettre en évidence que les écarts. Cet article examine comment des réseaux de neurones pourraient apprendre à effectuer ces comparaisons de façon fiable, et ce qui se passe quand cet apprentissage est perturbé.
Comment le cerveau prédit son propre monde sensoriel
Les scientifiques savent depuis longtemps que le cerveau envoie une copie interne des commandes motrices, appelée décharge corollaire, aux aires sensorielles. Ce signal porte une prédiction de ce que nous devrions voir ou entendre lorsque nous nous déplaçons. Si le stimulus entrant du monde extérieur correspond à cette prédiction, les réponses sensorielles sont atténuées ; sinon, certains neurones répondent fortement, signalant une erreur de prédiction. Les auteurs se concentrent sur deux types de neurones de ce type : ceux qui répondent lorsque le stimulus est plus fort que prévu (erreur de prédiction positive) et ceux qui répondent lorsque la prédiction est supérieure au stimulus réel (erreur de prédiction négative). Ensemble, ils permettent au cerveau de détecter à la fois les sensations manquantes et les sensations inattendues, comme un son qui ne se produit pas quand nous agissons, ou un son qui apparaît sans mouvement correspondant. 
Un signal d’enseignement global pour un câblage local
À première vue, calculer un écart semble simple : soustraire la prédiction du stimulus et prendre la valeur absolue. Mais les circuits corticaux réels affrontent plusieurs obstacles. Les connexions longue distance entre zones cérébrales sont majoritairement excitatrices, tandis que des cellules inhibitrices locales sont nécessaires pour annuler les entrées. De plus, les neurones du cortex sensoriel ont tendance à émettre à de faibles taux de base, ce qui complique la représentation des écarts positifs et négatifs. La proposition centrale de ce travail est une règle d’apprentissage « à trois facteurs » biologiquement plausible qui permet aux synapses inhibitrices locales de se rewiring en utilisant un seul signal d’enseignement global. Ce signal, dont on pense qu’il est véhiculé par des neuromodulateurs comme la noradrénaline provenant du tronc cérébral, reflète l’ampleur globale de l’écart, sans avoir besoin de savoir quelle synapse spécifique en est responsable.
Comment les circuits locaux apprennent à signaler la surprise
Dans le modèle, chaque neurone pyramidale reçoit à la fois des entrées stimulus et prédiction, ainsi qu’une inhibition acheminée via des interneurones locaux. Initialement, l’inhibition est à peu près uniforme et ne parvient pas à annuler les bonnes entrées, si bien que les événements attendus déclenchent encore de fortes réponses. Pendant l’apprentissage, lorsque prédiction et stimulus concordent habituellement, le signal global a un signe donné et induit un ajustement de type hebbien qui équilibre l’excitation et l’inhibition totales, poussant le système sur une « ligne d’apprentissage lente » où l’activité globale est faible dans les conditions attendues. Lors des essais plus rares de discordance, le signe du signal global s’inverse, et la même règle locale devient anti-hebbienne, poussant les poids synaptiques le long de cette ligne jusqu’à ce que chaque neurone devienne spécialisé. Un sous-groupe devient des neurones d’erreur de prédiction positive, excités principalement quand le stimulus dépasse la prédiction ; un autre devient des neurones d’erreur de prédiction négative, répondant lorsque la prédiction surpasse le stimulus. Les auteurs montrent mathématiquement que cette règle simple converge vers la même solution optimale que la descente de gradient, mais en n’utilisant que des ingrédients biologiquement réalistes. 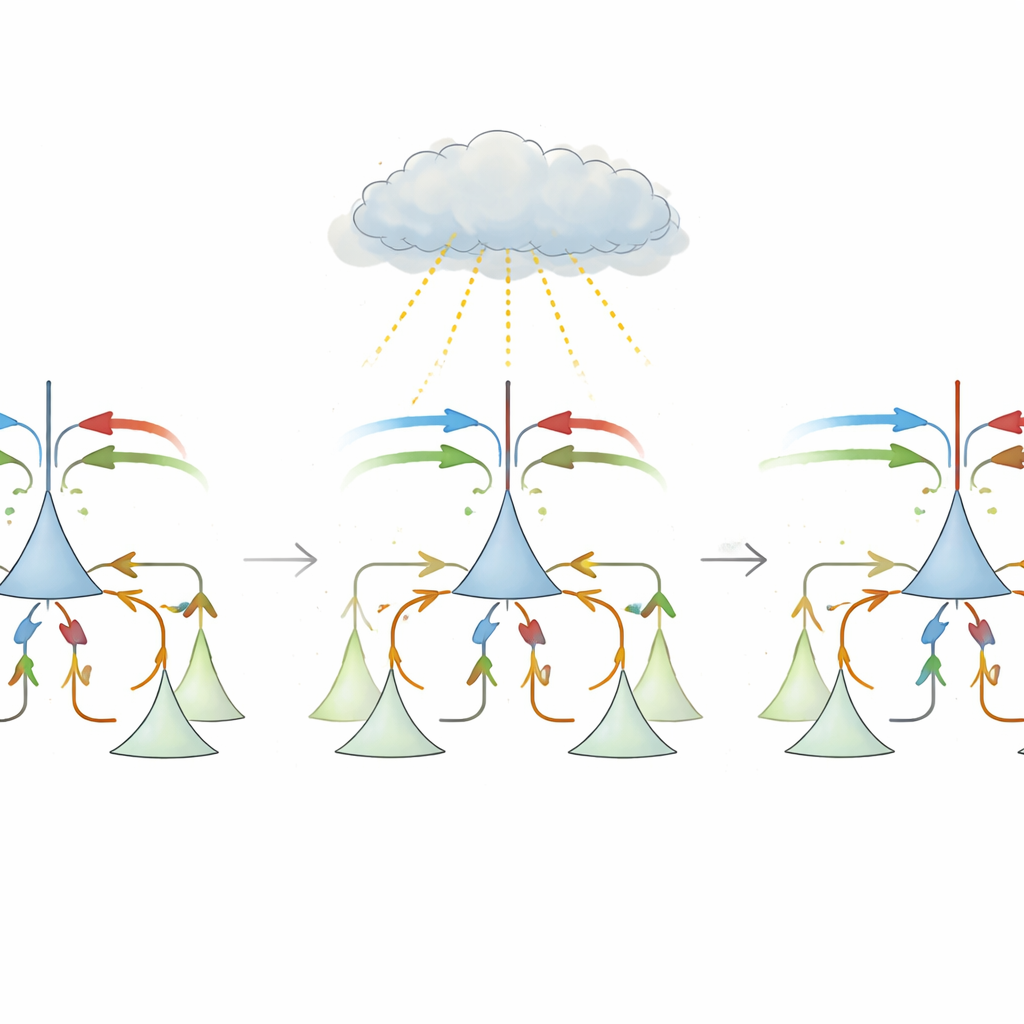
Relier le modèle aux données cérébrales réelles
Les chercheurs passent ensuite d’unités simplifiées à des modèles neuronaux plus réalistes et testent si les circuits appris reproduisent des expériences connues chez la souris. Dans un paradigme, les souris apprennent que courir sur un tapis entraîne un mouvement visuel ; plus tard, arrêter brièvement la scène visuelle pendant le mouvement provoque une forte réponse de discordance dans le cortex visuel. Le modèle montre le même schéma : après apprentissage, l’activité liée au mouvement attendu est supprimée, tandis que les arrêts soudains évoquent de fortes réponses, en particulier chez les neurones d’erreur de prédiction négative qui sont normalement freinés par l’entrée visuelle. Lorsque les auteurs simulent la perturbation de différentes classes de cellules inhibitrices, ou le blocage des récepteurs NMDA qui soutiennent la plasticité, la capacité du réseau à signaler les discordances se dégrade de façons qui correspondent aux observations pharmacologiques et optogénétiques. Enfin, le modèle prédit qu’après apprentissage, les neurones devraient se scinder en deux groupes avec des corrélations opposées à la force du stimulus et de la prédiction. Une réanalyse de données murines existantes confirme cela : seuls les animaux avec un appariement mouvement–vision correct montrent un schéma bimodal clair, cohérent avec des populations distinctes d’erreurs de prédiction positives et négatives.
Pourquoi cela compte pour la perception et la santé mentale
L’étude soutient une vue unificatrice selon laquelle une diffusion d’erreur globale à l’échelle du cerveau régule des connexions locales très spécifiques afin que les aires sensorielles deviennent expertes pour annuler le prévisible et mettre en valeur le surprenant. Dans la vie quotidienne, cela stabilise notre perception pendant nos propres mouvements tout en nous gardant sensibles aux événements inattendus. Le même mécanisme peut aussi alléger les régions cérébrales supérieures du travail de prédiction détaillée, libérant des ressources cognitives une fois les compétences apprises. Comme des perturbations de la décharge corollaire et du signalement d’erreur de prédiction ont été associées à des hallucinations et des délires, le mécanisme proposé offre aussi un cadre concret de circuit neural pour comprendre comment ces symptômes pourraient apparaître lorsque l’apprentissage inhibiteur ou les signaux d’erreur neuromodulateurs dysfonctionnent.
Citation: Meng, J.H., Wang, XJ. Global error signal guides local optimization in mismatch calculation. Nat Commun 17, 3868 (2026). https://doi.org/10.1038/s41467-026-70354-x
Mots-clés: codage prédictif, décharge corollaire, plasticité inhibitrice, neuromodulation, neurones d’erreur de prédiction