Clear Sky Science · en
Global error signal guides local optimization in mismatch calculation
Why our brains don’t startle at our own actions
When you click a mouse to play a song, you are not surprised by the sound that follows. But if music suddenly starts without your touch, it grabs your attention. This everyday difference between expected and unexpected sensations relies on the brain’s ability to predict the sensory consequences of our own movements and to highlight only the mismatches. This paper explores how networks of neurons might learn to make those comparisons reliably, and what goes wrong when that learning is disrupted.
How the brain predicts its own sensory world
Scientists have long known that the brain sends an internal copy of movement commands, called a corollary discharge, to sensory areas. This signal carries a prediction of what we should see or hear when we move. If the incoming stimulus from the outside world matches this prediction, sensory responses are dampened; if it does not, certain neurons respond strongly, signaling a prediction error. The authors focus on two kinds of such neurons: ones that respond when the stimulus is stronger than predicted (positive prediction error), and ones that respond when the prediction is stronger than the actual stimulus (negative prediction error). Together, they allow the brain to detect both missing and unexpected sensations, such as a sound that fails to occur when we act, or a sound that appears without a matching movement. 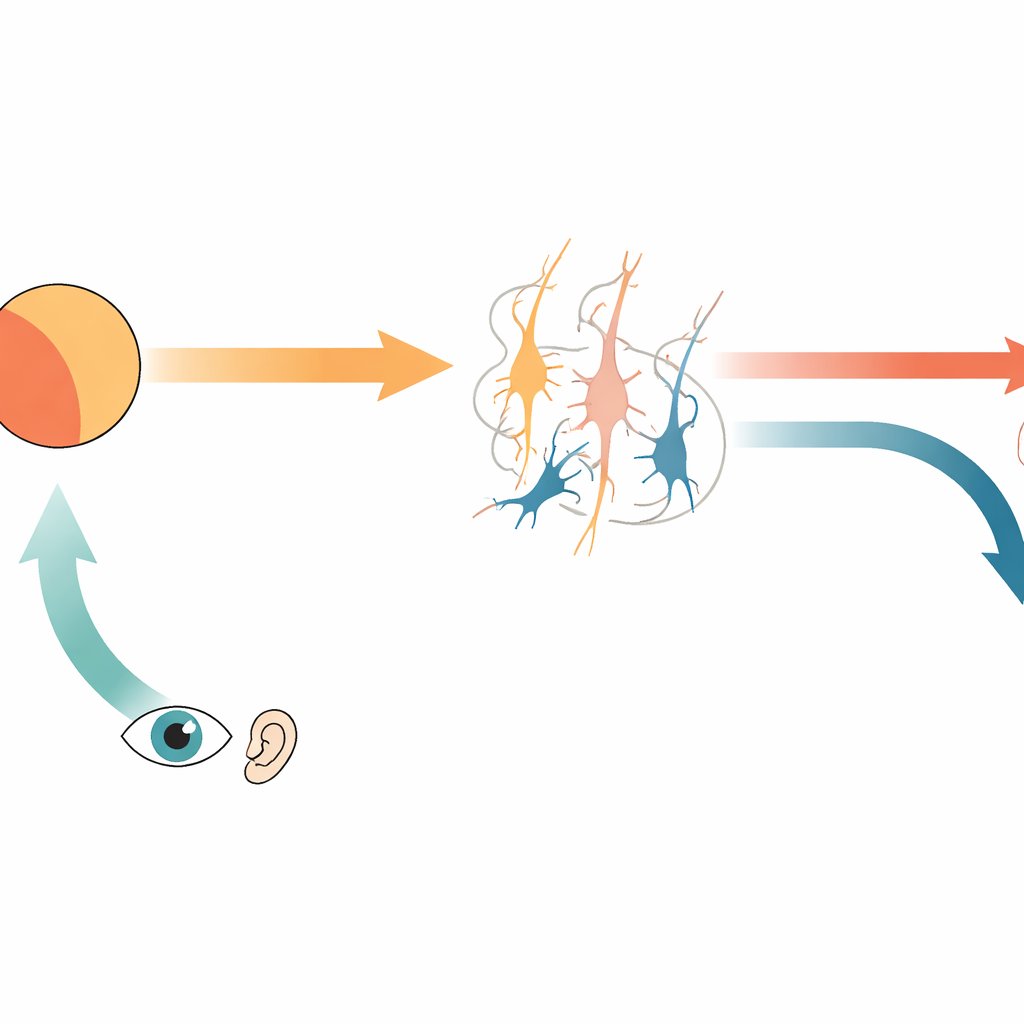
A global teaching signal for local wiring
At first glance, computing a mismatch seems simple: subtract prediction from stimulus and take the absolute value. But real cortical circuits face several hurdles. Long-range connections between brain areas are mostly excitatory, while local inhibitory cells are needed to cancel inputs. In addition, neurons in sensory cortex tend to fire at low baseline rates, which complicates representing both positive and negative deviations. The central proposal of this work is a biologically plausible “three-factor” learning rule that lets local inhibitory synapses rewire themselves using a single global teaching signal. This signal, thought to be carried by neuromodulators such as noradrenaline from the brainstem, reflects how big the mismatch is overall, without needing to know which specific synapse caused it.
How local circuits learn to signal surprise
In the model, each pyramidal neuron receives both stimulus and prediction inputs, as well as inhibition routed through local interneurons. Initially, inhibition is roughly uniform and fails to cancel the right inputs, so expected events still evoke strong responses. During learning, when prediction and stimulus usually match, the global signal has one sign and drives a Hebbian-like adjustment that balances total excitation and inhibition, pushing the system onto a “slow learning” line where overall activity is low in expected conditions. On rarer mismatch trials, the sign of the global signal flips, and the same local rule becomes anti-Hebbian, nudging synaptic weights along that line until each neuron ends up specialized. One subgroup becomes positive prediction error neurons, excited mainly when stimulus exceeds prediction; another becomes negative prediction error neurons, responding when prediction overshoots the stimulus. The authors show mathematically that this simple rule converges to the same optimal solution as gradient descent, but using only biologically realistic ingredients. 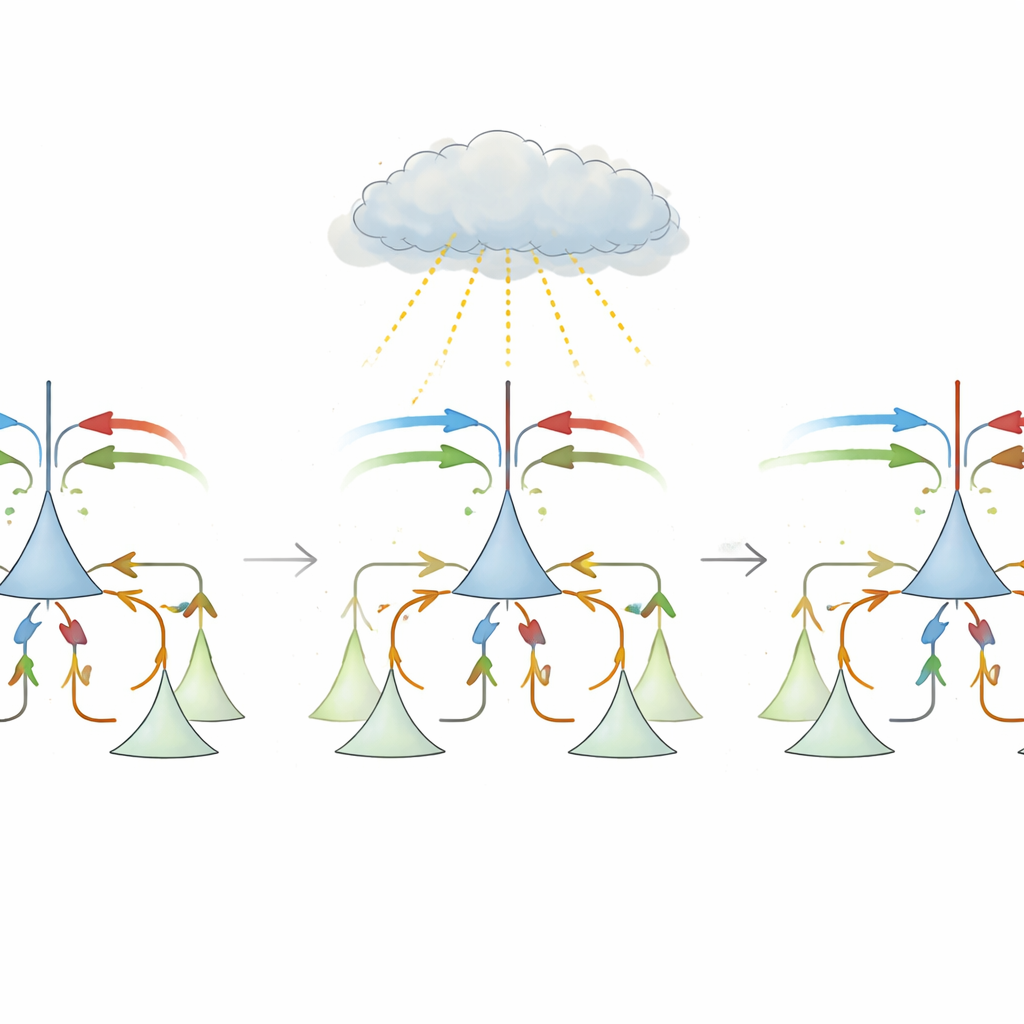
Linking the model to real brain data
The researchers then move from simplified units to more realistic neuron models and test whether the learned circuits reproduce known experiments in mice. In one paradigm, mice learn that running on a treadmill causes visual motion; later, briefly pausing the visual scene during movement causes a strong mismatch response in visual cortex. The model shows the same pattern: after learning, activity for expected motion is suppressed, whereas sudden pauses evoke large responses, especially in negative prediction error neurons that are normally held back by visual input. When the authors simulate disturbing different classes of inhibitory cells, or blocking NMDA receptors that support plasticity, the network’s ability to signal mismatch degrades in ways that align with pharmacological and optogenetic findings. Finally, the model predicts that, after learning, neurons should split into two groups with opposite correlations to stimulus and prediction strength. Reanalysis of existing mouse data confirms this: only animals with proper movement–vision coupling show a clear bimodal pattern, consistent with distinct positive and negative prediction error populations.
Why this matters for perception and mental health
The study supports a unifying view in which a global brain-wide error broadcast tunes very specific local connections so that sensory areas become expert at cancelling the predictable and highlighting the surprising. In everyday life, this makes our perception stable during our own movements while keeping us sensitive to unexpected events. The same machinery may also help relieve higher brain regions of detailed prediction work, freeing cognitive resources once skills are learned. Because disruptions of corollary discharge and prediction error signaling have been linked to hallucinations and delusions, the proposed mechanism also offers a concrete neural circuit framework for understanding how such symptoms might arise when inhibitory learning or neuromodulatory error signals go awry.
Citation: Meng, J.H., Wang, XJ. Global error signal guides local optimization in mismatch calculation. Nat Commun 17, 3868 (2026). https://doi.org/10.1038/s41467-026-70354-x
Keywords: predictive coding, corollary discharge, inhibitory plasticity, neuromodulation, prediction error neurons