Clear Sky Science · pl
Mechanizmy rozwojowe niezależne od actinotrichia u kolczastych promieni ułatwiają morfologiczną dywersyfikację płetw ryb Acanthomorpha
Dlaczego „kręgosłupy” płetw mają znaczenie dla różnorodności ryb
Od latających ryb szybujących nad falami po węgorzycowate zwabiające zdobycz świecącymi przynętami — wiele z najbardziej niezwykłych przystosowań ryb opiera się na jednej kluczowej strukturze: sztywnych kolcach płetwowych. W tym badaniu postawiono na pozór proste pytanie o duże konsekwencje ewolucyjne: czym różni się sposób wzrostu tych kolców od zwykłych, giętkich promieni płetwowych? Odkrywając ten ukryty plan budowy, autorzy pokazują, jak niewielkie zmiany w komórkach i materiałach podporowych mogą otworzyć drogę do ogromnej różnorodności form.
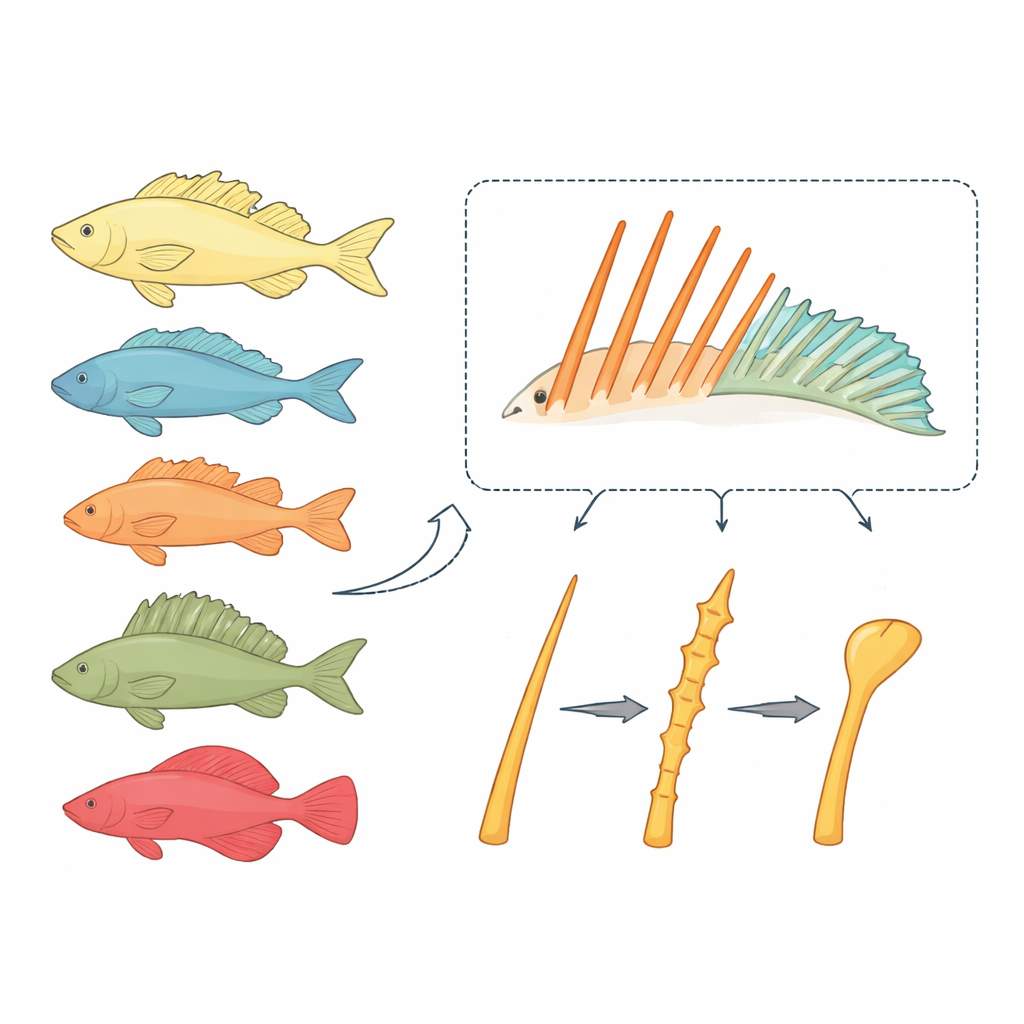
Dwa rodzaje „prętów” płetw o bardzo różnych losach
Ryby kostnoszkieletowe zwykle mają dwa typy podpór płetw: miękkie promienie, które wyginają się i rozgałęziają niczym drobne palce, oraz kolczaste promienie — sztywne, ostre pręty. Miękkie promienie rzadko zmieniają swoją podstawową konstrukcję prętową, nawet gdy osiągają duże rozmiary. W przeciwieństwie do nich kolczaste promienie w dużej grupie ryb Acanthomorpha były wielokrotnie przekształcane w nowe „narzędzia”, w tym dysk przyssawkowy remory czy wysuwany „wędkowy” kolc u węgorzycowatych. Badacze podejrzewali, że rozwój kolców podąża własnymi zasadami i że te zasady mogą wyjaśniać, dlaczego kolce są tak elastyczne ewolucyjnie.
Nowy gatunek laboratoryjny do obserwacji wzrostu kolców
Tradycyjne modele, takie jak Danio pręgowany (zebrafish) czy medaka, słabo nadają się do badania prawdziwych kolczastych promieni: zebrafish ich nie mają, a medaka ma tylko szczątkowy kolec. Zespół zwrócił się zamiast tego ku karłowatej neonowej tęczance, Melanotaenia praecox — małej słodkowodnej rybie, która rozwija wyraźne zestawy zarówno kolczastych, jak i miękkich promieni i nadaje się do manipulacji genetycznych. Barwiąc rosnące kości w dwóch punktach czasowych, wykazali, że kolczaste promienie u tęczanki wydłużają się przez dodawanie nowej kości na końcach, podobnie jak promienie miękkie. Jednak kolce także pogrubiają się, gdy pary lewe i prawe połówek zrastają się w pojedynczy, sztywny słup — cechę charakterystyczną tych struktur.
Rezygnacja z kolagenowych rusztowań na rzecz komórkowych „czapek”
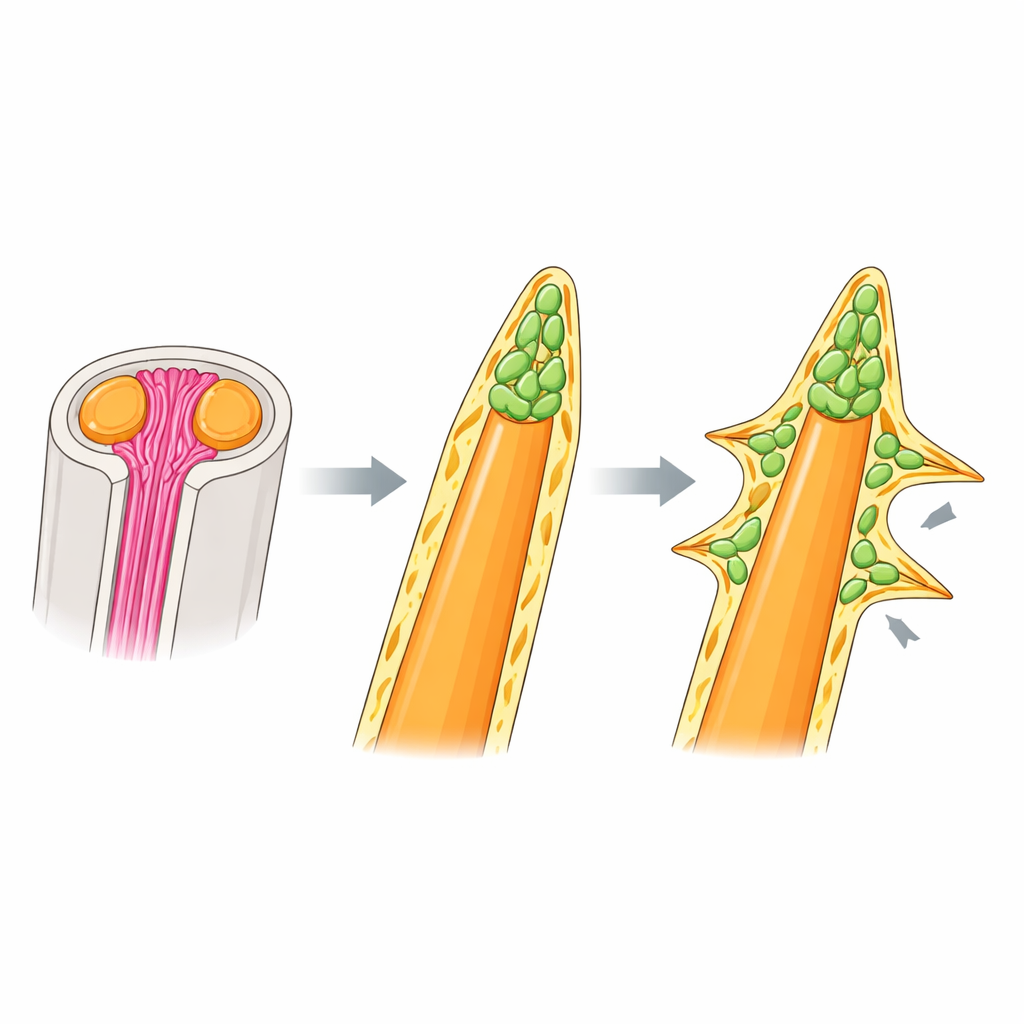
Wiadomo, że miękkie promienie u innych ryb opierają się na igłopodobnych włóknach kolagenowych, zwanych actinotrichia, które zbierają się na końcach promieni i prowadzą komórki tworzące kość. Ku zaskoczeniu autorów, kolczaste promienie tęczanki nie używają tego rusztowania. Fluorescencyjne barwienia kolagenu oraz ekspresja kluczowych genów actinotrichia były wyraźne na końcach promieni miękkich, lecz słabe lub nieobecne wokół kolców, a mutanty pozbawione actinotrichia miały skręcone, zdeformowane promienie miękkie, podczas gdy kolczaste pozostały całkowicie normalne. Mikroskopia elektronowa i obrazowanie 3D ujawniły, co stosują kolce zamiast tego: gęstą czapę z komórek mezenchymalnych na końcu kości, owiniętą grubą, podobną do błony podstawnej warstwą macierzy zewnątrzkomórkowej. Ta czapa znajduje się tam, gdzie w promieniach miękkich byłyby actinotrichia, i wydaje się napędzać zarówno wydłużanie, jak i pogrubianie kolca niezależnie od pakietów kolagenu.
Sygnalizacja rzeźbiąca kolce i inne kształty
W obrębie komórkowej czapy na końcach kolców tęczanki wiele komórek nosiło markery niedojrzałych komórek tworzących kość (osteoblastów) i wykazywało aktywność sygnalizacji BMP — szlaku znanego z promowania różnicowania kości. Kiedy zespół zablokował receptory BMP lekiem, kolce nie przekształciły się w promienie miękkie, lecz ich końce stały się bardziej tępe i nieprawidłowo pogrubione, z uwięzionymi wewnątrz kości komórkami. Wskazuje to, że sygnalizacja BMP precyzuje, ile kości jest dodawane i jak porządnie jest układana, zamiast decydować, czy promień będzie kolczasty czy miękki. Aby przetestować, czy manipulacja tym samym mechanizmem może wygenerować egzotyczne kształty kolców, autorzy zbadali larwy mola piłokształtnego, którego grzbietowy kolec nosi rząd kolczastych wyrostków bocznych. Tam również skupiska niedojrzałych osteoblastów z aktywną sygnalizacją BMP pojawiały się nie tylko na głównym wierzchołku, lecz także przy każdym bocznym wyrostku, sugerując, że podstawowy program „czapy wierzchołkowej” został powtórnie użyty i przemieszczo ny, by zbudować kolce boczne.
Jak elastyczne zasady budowy napędzają ewolucyjne innowacje
W sumie praca przedstawia kolczaste promienie jako struktury budowane przez mobilne komórki tworzące kość, a nie związane z ustalonymi kolagenowymi przewodnikami. Ponieważ ich wzrost nie jest zakotwiczony w prostych pakietach actinotrichia, kierunek i miejsce rozrostu kolca mogą się zmieniać, gdy tylko skupiska osteoblastów i ogniska sygnalizacyjne przesuną się wzdłuż kości. Autorzy przekonują, że ta rozwojowa swoboda ułatwiła ewolucji przekształcanie prostych kolców płetwowych w różnorodne, czasem dziwaczne „narzędzia” obserwowane u acanthomorficznych ryb. Szerzej, badanie podkreśla, jak zmiany w tym, gdzie komórki się gromadzą i jakich materiałów zewnątrzkomórkowych używają — bez modyfikowania podstawowych zestawów genetycznych — mogą napędzać istotne innowacje kształtu ciała u zwierząt.
Cytowanie: Miyamoto, K., Kuroda, J., Kamimura, S. et al. Actinotrichia-independent developmental mechanisms of spiny rays facilitate the morphological diversification of Acanthomorpha fish fins. Nat Commun 17, 2775 (2026). https://doi.org/10.1038/s41467-026-69180-y
Słowa kluczowe: ewolucja płetw ryb, kolczaste promienie, rozwój kości, sygnalizacja osteoblastów, morficzna dywersyfikacja