Clear Sky Science · de
Actinotrichia-unabhängige Entwicklungsmechanismen von Stachelrippen fördern die morphologische Diversifizierung der Flossen bei Acanthomorpha-Fischen
Warum Flossenachsen für die Fischvielfalt wichtig sind
Von fliegenden Fischen, die über die Wellen gleiten, bis zu Anglerfischen mit leuchtenden Ködern — viele der seltsamsten Fischtricks beruhen auf einer Schlüsselkonstruktion: starren Flossenstacheln. Die vorliegende Studie stellt eine auf den ersten Blick einfache, evolutionär aber weitreichende Frage: Was unterscheidet das Wachstum dieser Stacheln von dem gewöhnlicher flexibler Flossenstrahlen? Indem die Autoren diesen verborgenen Bauplan aufdecken, zeigen sie, wie kleine Veränderungen in Zellen und Stützmaterialien eine enorme Vielfalt an Formen möglich machen können.
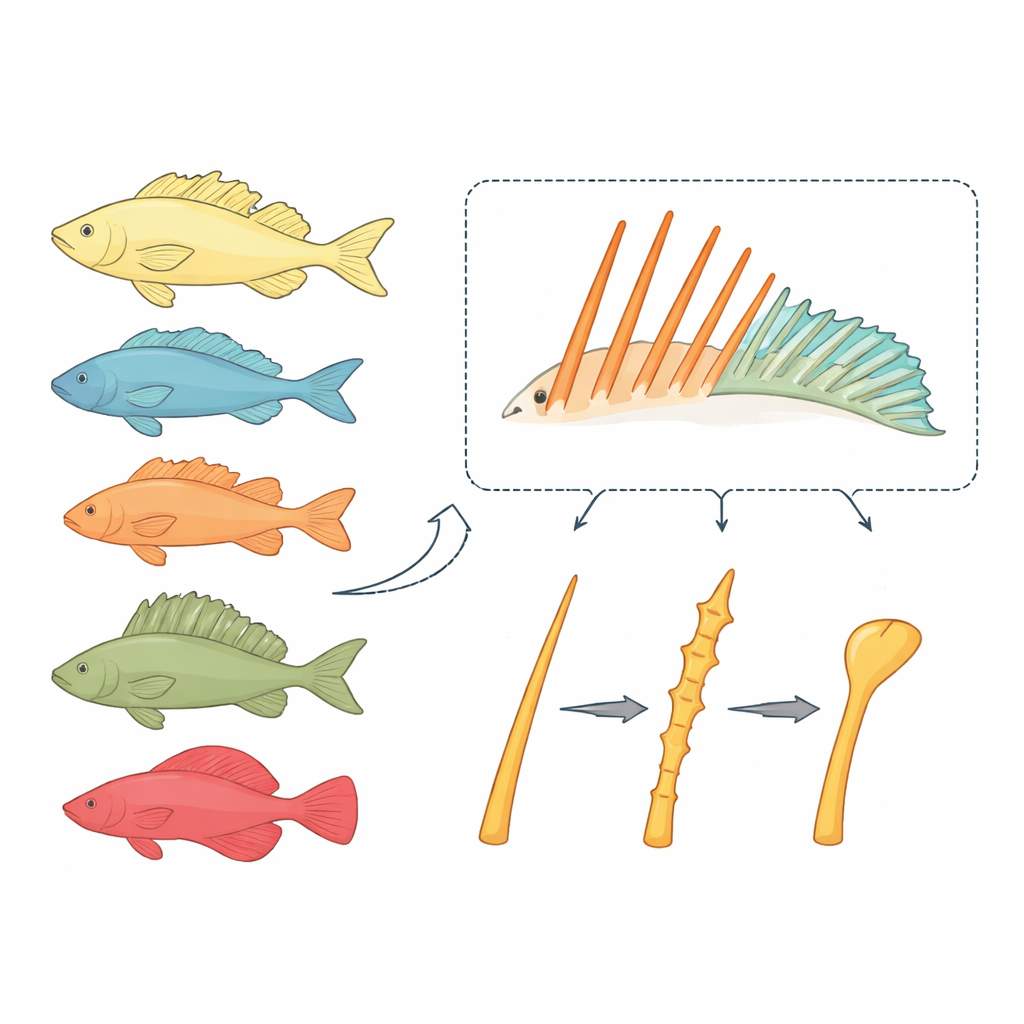
Zwei Arten von Flossenstäben mit sehr unterschiedlichen Schicksalen
Knochenfische tragen typischerweise zwei Typen von Flossenstützen: weiche Strahlen, die sich biegen und verzweigen wie kleine Finger, und Stachelrippen, die steif, spitz und stabförmig sind. Weiche Strahlen behalten selten ihr grundlegendes stabähnliches Design bei, selbst wenn sie sehr lang werden. Im Gegensatz dazu wurden Stachelrippen in der großen Fischgruppe der Acanthomorpha wiederholt zu neuartigen „Gadgets“ umgestaltet, etwa zur Saugscheibe der Remora oder zur Angelrute der Anglerfische. Die Forscher vermuteten, dass die Entwicklung von Stachelrippen eigenen Regeln folgt und dass diese Regeln erklären könnten, warum Stacheln evolutionsbiologisch so flexibel sind.
Ein neues Laborfisch-Modell, um Stacheln beim Wachsen zu beobachten
Traditionelle Modellarten wie Zebrafisch und Medaka eignen sich schlecht, um echte Stachelrippen zu untersuchen: Zebrafische haben sie nicht, und Medaka besitzt nur einen rudimentären Stachel. Das Team wandte sich stattdessen dem Zwerg-Neonregenbogenfisch, Melanotaenia praecox, zu — einer kleinen Süßwasserart, die deutlich ausgeprägte Sätze sowohl von stacheligen als auch von weichen Strahlen entwickelt und genetisch manipulierbar ist. Durch Färben wachsender Knochen zu zwei Zeitpunkten zeigten sie, dass sich Regenbogenfisch-Stachelrippen durch Einlagerung neuen Knochens an den Spitzen verlängern, ähnlich wie weiche Strahlen. Stacheln verdicken sich jedoch zusätzlich, wenn die paarigen linken und rechten Hälften zu einer starren Säule verschmelzen — ein Kennzeichen dieser Strukturen.
Loslassen der Kollagen-Gerüste und stattdessen Zellkappen verwenden
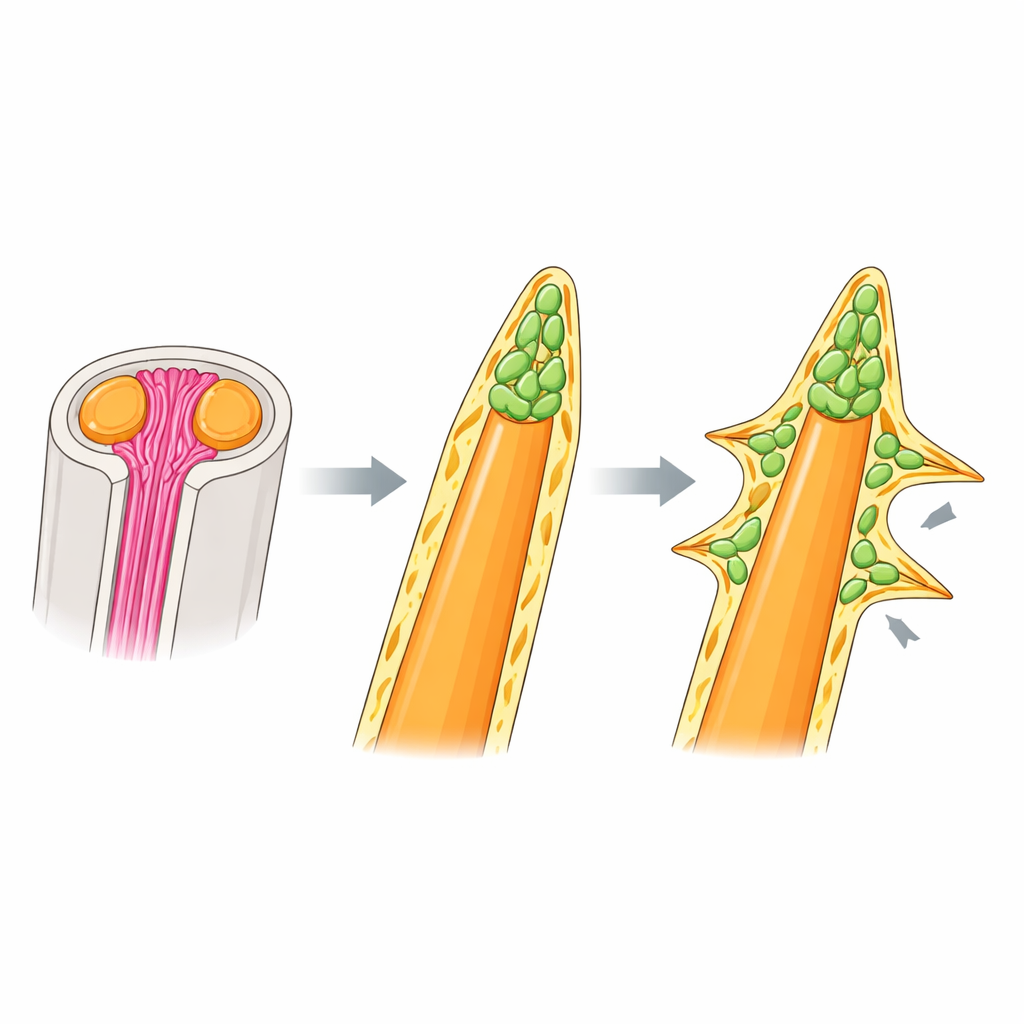
Bei weichen Strahlen anderer Fische ist bekannt, dass sie auf nadelartigen Kollagenfasern, den Actinotrichia, beruhen, die sich an den Strahlspitzen bündeln und knochenbildende Zellen führen. Überraschenderweise fanden die Autoren, dass Regenbogenfisch-Stachelrippen dieses Gerüst nicht nutzen. Fluoreszierende Kollagenfärbungen und die Expression wichtiger Actinotrichia-Gene zeigten starke Signale an den Spitzen weicher Strahlen, waren aber um Stacheln schwach oder fehlten. Mutanten ohne Actinotrichia wiesen eingerollte, missgestaltete weiche Strahlen, aber völlig normale Stachelrippen auf. Elektronenmikroskopie und 3D-Bildgebung enthüllten, was Stacheln stattdessen verwenden: eine dichte Kappe aus mesenchymalen Zellen an der Knochenspitze, umhüllt von einer dicken, basalmembranähnlichen Schicht der extrazellulären Matrix. Diese Kappe sitzt dort, wo bei weichen Strahlen die Actinotrichia wären, und scheint sowohl die Verlängerung als auch die Verdickung des Stachels unabhängig von Kollagenbündeln zu steuern.
Signalgebung, die Dornen und andere Stachelformen formt
Innerhalb der Zellkappe an den Stachelspitzen des Regenbogenfischs trugen viele Zellen Marker unreifer knochenbildender Zellen (Osteoblasten) und zeigten Aktivität der BMP-Signalgebung — eines Weges, der bekanntermaßen die Knochenbildung fördert. Als das Team BMP-Rezeptoren mit einem Wirkstoff blockierte, verwandelten sich die Stacheln nicht in weiche Strahlen, aber ihre Spitzen wurden stumpfer und ungewöhnlich dick, mit Zellen, die im Knochen eingeschlossen waren. Das deutet darauf hin, dass BMP-Signalgebung eher feinabstimmt, wie viel Knochen hinzugefügt und wie ordentlich er eingelagert wird, statt zu bestimmen, ob ein Strahl stachelig oder weich wird. Um zu prüfen, ob das gleiche System exotische Stachelformen erzeugen kann, untersuchten die Autoren Larven eines Feilenfischs, dessen dorsaler Stachel eine Reihe dorniger Seitenfortsätze trägt. Auch dort erschienen Cluster unreifer Osteoblasten mit aktiver BMP-Signalgebung nicht nur an der Hauptspitze, sondern an jeder lateralen Ausstülpung, was darauf hindeutet, dass das grundlegende "Spitzenkappen"-Programm wiederverwendet und neu positioniert wurde, um Dornen zu bauen.
Wie flexible Bauregeln evolutionäre Erfindung antreiben
Insgesamt zeichnen die Ergebnisse Stachelrippen als Strukturen, die von mobilen knochenbildenden Zellen gebaut werden, statt an feste Kollagenschienen gebunden zu sein. Weil ihr Wachstum nicht an gerade Actinotrichia-Bündel gekoppelt ist, können Richtung und Ort der Stachelerweiterung sich verändern, sobald sich Osteoblast-Cluster und Signalisierungs-Hotspots entlang des Knochens verschieben. Die Autoren argumentieren, dass diese entwicklungsbiologische Freiheit es der Evolution erleichtert hat, einfache Flossenstacheln in die vielfältigen, mitunter bizarren Apparate zu verwandeln, die bei Acanthomorpha-Fischen vorkommen. Allgemeiner hebt die Studie hervor, wie Änderungen darin, wo Zellen sich ansammeln und welche extrazellulären Materialien sie verwenden — ohne die grundlegenden genetischen Werkzeuge zu verändern — große Innovationen in der Körperform über Tierarten hinweg antreiben können.
Zitation: Miyamoto, K., Kuroda, J., Kamimura, S. et al. Actinotrichia-independent developmental mechanisms of spiny rays facilitate the morphological diversification of Acanthomorpha fish fins. Nat Commun 17, 2775 (2026). https://doi.org/10.1038/s41467-026-69180-y
Schlüsselwörter: Evolution der Fischflosse, Stachelrippen, Knochenentwicklung, Osteoblastensignalisierung, Morphologische Diversifizierung