Clear Sky Science · it
Vescicola otica di zebrafish ed epididimo murino come modelli per studiare la divisione delle cellule epiteliali colonnari
Come i nostri tessuti si dividono senza disfarsi
Ogni momento, le cellule che rivestono i nostri organi si dividono, sostituendo vicini logori mantenendo intatti e funzionanti tessuti come l’intestino, l’orecchio interno e le vie riproduttive. Se questa coreografia fallisce, le conseguenze possono includere difetti congeniti, infertilità e cancro. Questo studio presenta due potenti modelli viventi — un piccolo orecchio di pesce e un dotto riproduttivo di topo — per osservare, in tempo reale, come cellule epiteliali alte e compatte si dividono senza rompere la barriera che formano.
Osservare i nuclei cellulari compiere un pendolarismo quotidiano
In molti organi, le cellule che formano fogli sottili sono alte e a forma di colonna, con un’estremità attaccata a uno strato di sostegno e l’altra rivolta verso uno spazio pieno di fluido. Prima di dividersi, i loro nuclei compiono un “pendolarismo” attentamente temporizzato. Partono vicino alla base, si spostano verso la superficie interna, si dividono lì e poi i due nuovi nuclei tornano indietro. Questo viaggio avanti e indietro, chiamato migrazione nucleare interchinetica, è stato studiato soprattutto nel tessuto cerebrale in sviluppo. Gli autori hanno ragionato che per capire davvero come funziona questo processo nel corpo servivano osservazioni viventi ad alta risoluzione in tessuti più semplici ma ancora realistici.
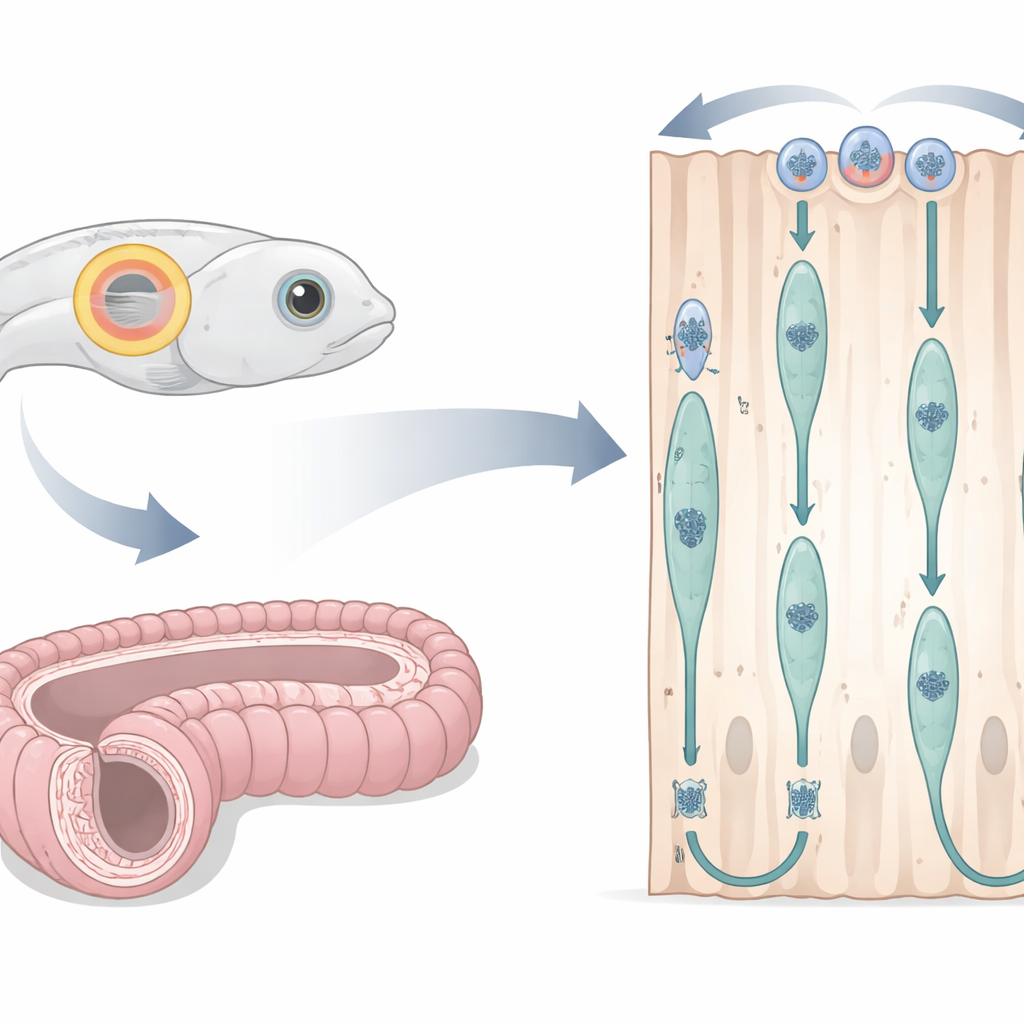
Un orecchio di pesce e un dotto di topo come laboratori viventi
I ricercatori hanno adattato la vescicola otica embrionale del zebrafish — una sacca trasparente piena di fluido che diventerà l’orecchio interno — come finestra sulla divisione cellulare in epitelio colonnare semplice. Iniettando marcatori fluorescenti nelle uova fecondate, hanno potuto vedere simultaneamente i contorni cellulari, i nuclei e fibre strutturali chiave al microscopio confocale mentre l’embrione si sviluppava. Si sono concentrati su una finestra temporale in cui le cellule formavano un anello ordinato e alto intorno alla vescicola. Parallelamente, hanno esaminato l’epididimo del topo, un lungo tubo avvolto che aiuta la maturazione degli spermatozoi, usando sezioni di tessuto sottili e marcatori chimici che evidenziano la sintesi del DNA, le fasi del ciclo cellulare e le proteine strutturali. Questo disegno cross-specie ha permesso di testare se lo stesso ballo nucleare si svolga in vertebrati molto diversi.
Le forze che tirano e modellano le cellule in divisione
Un tracciamento accurato nel zebrafish ha rivelato che il nucleo inizia il suo movimento verso l’alto solo nella fase tardiva della preparazione alla divisione, completando il viaggio in poco più di un’ora. Esperimenti che interferivano con diversi “motori” interni hanno mostrato che questo movimento dipende fortemente dai microtubuli — piste proteiche rigide che corrono dalla base alla punta — e dalla dineina, una proteina motrice che cammina su queste piste verso l’apice della cellula. Quando i microtubuli sono stati degradati con farmaci, la migrazione nucleare si è quasi fermata; quando l’attività della dineina è stata bloccata, molti meno nuclei hanno raggiunto la sommità. Al contrario, disabilitare la miosina II, il principale motore della contrazione basata su actina, non ha rallentato il movimento nucleare verso l’alto, rivelando che in questi epiteli alti ma relativamente semplici, il traino basato sui microtubuli è il motore chiave per posizionare il nucleo.
Arrotondarsi senza staccarsi
Quando il nucleo arriva vicino alla superficie interna, la cellula subisce un drammatico cambiamento di forma: si arrotonda per dividersi. Imaging vivo e colorazioni proteiche in zebrafish e topo hanno mostrato che durante questo “arrotondamento” il corpo principale della cellula si gonfia verso il lume mentre un sottile stelo di membrana la mantiene ancorata alla base. Actina e miosina II si concentrano lungo i lati della cellula, stringendoli come un laccio. Allo stesso tempo, le proteine che rafforzano le giunzioni cellula-cellula diventano più attive, aiutando la cellula in divisione a restare saldamente attaccata ai vicini. Quando la miosina II è stata inibita, le cellule non si arrotondavano correttamente, i loro fusi spesso si inclinavano rispetto al normale piano di divisione piatto e uno dei due nuclei figlio rimaneva frequentemente bloccato vicino alla superficie interna invece di reintegrarsi nel foglio. Questi risultati mostrano che, sebbene la miosina II non sia necessaria per spostare il nucleo verso l’alto, è cruciale per modellare la cellula in divisione e mantenere l’organizzazione tissutale.
La posizione come semaforo per la divisione
Lo studio ha inoltre scoperto un legame stretto tra la posizione del nucleo e la possibilità che la cellula entri nella fase finale della divisione. Nelle alte cellule dell’orecchio del zebrafish, i nuclei raramente iniziavano la mitosi a meno che non fossero prima arrivati vicino alla superficie interna. Quando i microtubuli sono stati disturbati, le cellule nelle regioni più spesse del tessuto per lo più si bloccavano prima della divisione, mentre le cellule nelle regioni più sottili — dove i nuclei erano già vicini alla sommità — potevano procedere. Nell’epididimo murino, piccole strutture chiamate centrioli restavano vicino alla superficie interna e solo quando il nucleo le raggiungeva i principali regolatori della divisione si spostavano nel nucleo e la macchina della divisione si assemblava. Questo suggerisce che, in questi epiteli, la posizione nucleare funge da guardiano: solo i nuclei che hanno completato con successo il loro viaggio verso l’alto ricevono i segnali necessari per impegnarsi nella divisione.
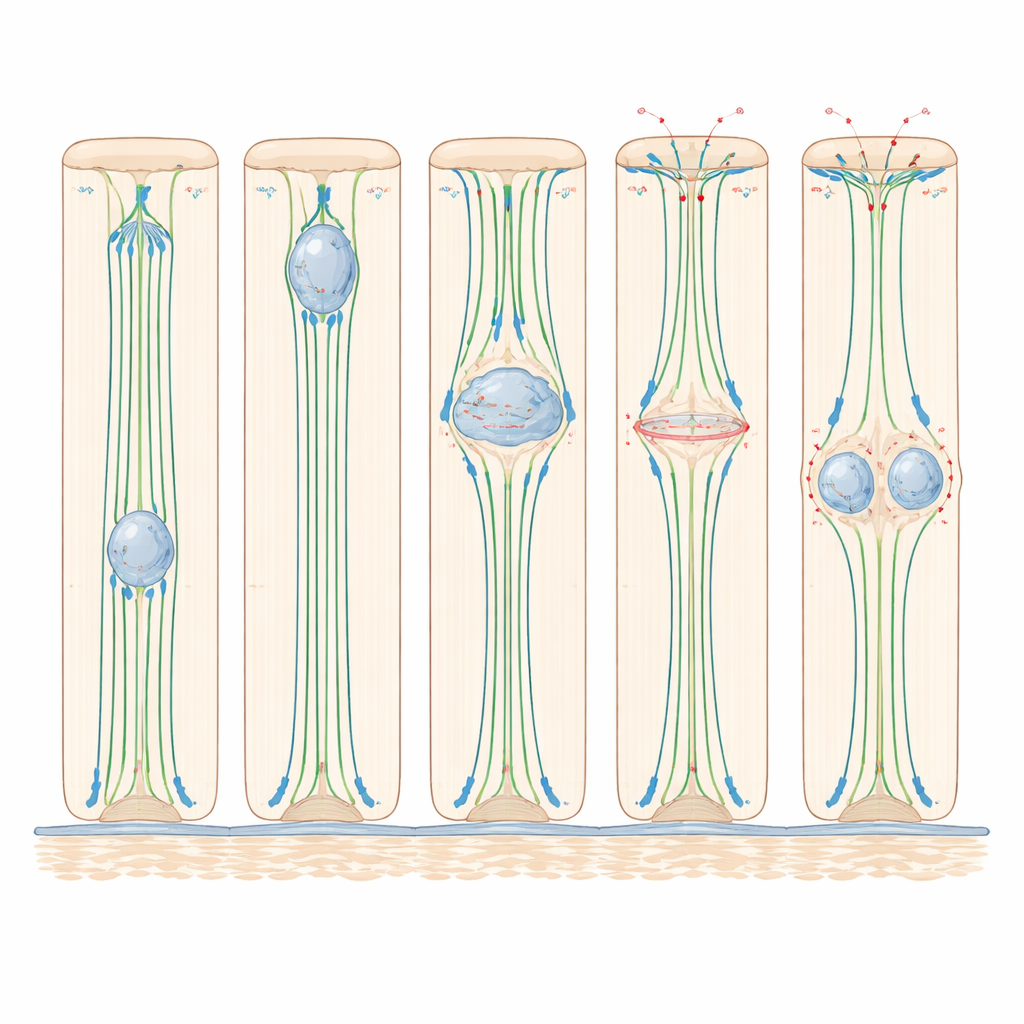
Cosa significa per tessuti sani
Nel complesso, il lavoro mostra che gli epiteli colonnari semplici di pesci e mammiferi condividono una strategia comune: microtubuli e dineina tirano i nuclei verso la superficie interna nelle fasi tardive del ciclo cellulare; l’arrotondamento guidato dall’actomiosina modella la cellula e orienta la divisione; e mantenere un sottile ancoraggio basale più un angolo di divisione piatto aiuta entrambe le cellule figlie a rientrare nello strato senza strapparlo. Stabilendo la vescicola otica del zebrafish e l’epididimo del topo come modelli complementari, lo studio fornisce una visione chiara e in azione di come le cellule in divisione salvaguardino la struttura tissutale — una base per ricerche future su come questi processi si guastino nelle malattie e su come possano essere sfruttati per la riparazione dei tessuti.
Citazione: Xia, Y., Perder, B., Yao, A.G.C. et al. Zebrafish otic vesicle and mouse epididymis as model systems for studying columnar epithelial cell division. Sci Rep 16, 12995 (2026). https://doi.org/10.1038/s41598-026-42729-z
Parole chiave: divisione delle cellule epiteliali, migrazione nucleare, modello zebrafish, epididimo murino, imaging cellulare