Clear Sky Science · es
Vesícula ótica de pez cebra y epidídimo de ratón como sistemas modelo para estudiar la división de células epiteliales columnares
Cómo se dividen nuestros tejidos sin desmoronarse
Cada momento, las células que recubren nuestros órganos se dividen, reemplazando a vecinas agotadas mientras mantienen tejidos como el intestino, el oído interno y el tracto reproductor intactos y funcionando. Si esta coreografía falla, las consecuencias pueden incluir defectos congénitos, infertilidad y cáncer. Este estudio presenta dos poderosos modelos vivos —una pequeña vesícula del oído de pez y un conducto reproductor de ratón— para observar, en tiempo real, cómo las células epiteliales altas y densamente empaquetadas se dividen sin romper la barrera que forman.
Observar los núcleos celulares en su viaje cotidiano
En muchos órganos, las células que forman láminas delgadas son altas y de forma columnar, con un extremo adherido a una capa de soporte y el otro orientado hacia un espacio lleno de fluido. Antes de dividirse, sus núcleos realizan un “viaje” cuidadosamente cronometrado. Comienzan cerca de la base, se desplazan hacia la superficie interna, se dividen allí y luego los dos núcleos nuevos regresan hacia abajo. Este idas y venidas, llamado migración nuclear interquinética, se ha estudiado principalmente en el tejido cerebral en desarrollo. Los autores razonaron que, para entender realmente cómo funciona este proceso en todo el organismo, necesitaban vistas en vivo y de alta resolución en tejidos más simples pero aún realistas.
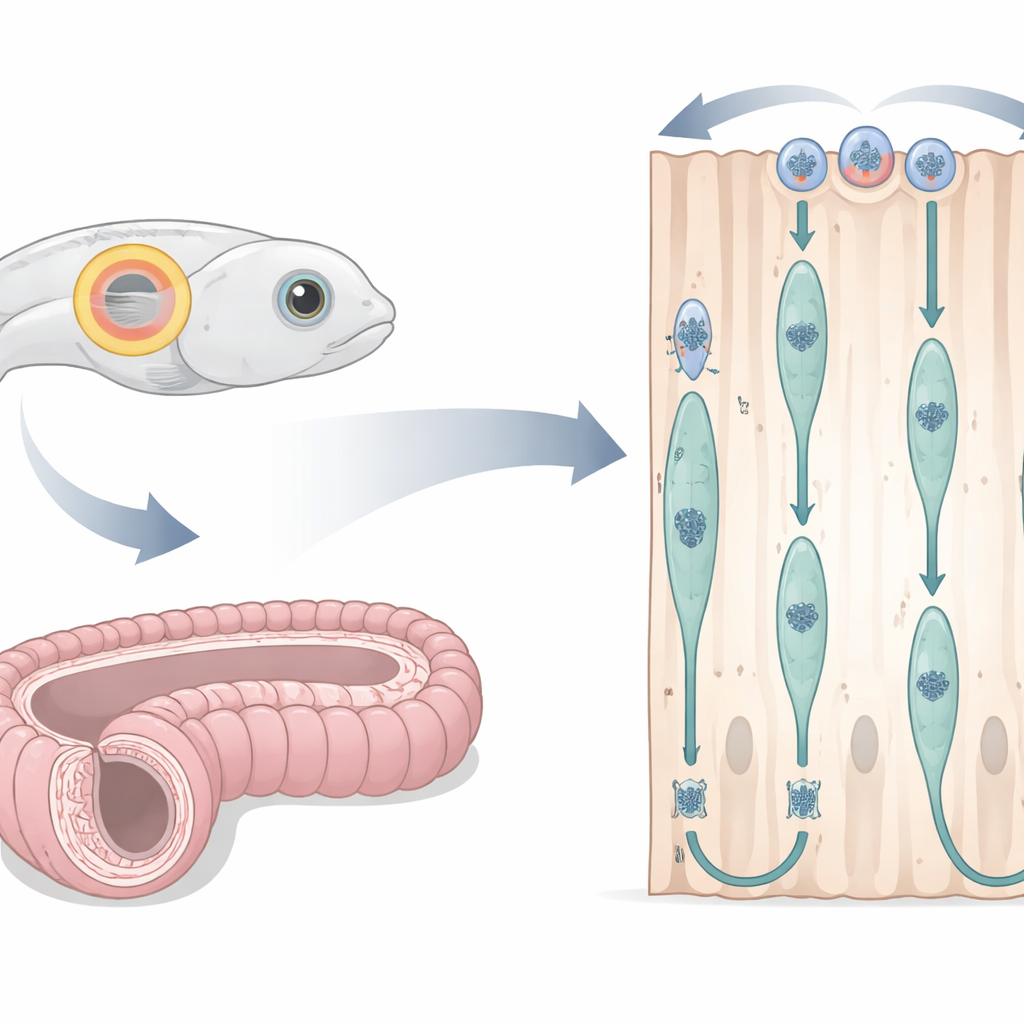
El oído de un pez y un conducto de ratón como laboratorios vivos
Los investigadores adaptaron la vesícula ótica embrionaria del pez cebra —un saco transparente y lleno de fluido que dará lugar al oído interno— como una ventana hacia la división celular en epitelio columnar simple. Al inyectar marcadores fluorescentes en los huevos fertilizados, pudieron ver simultáneamente los contornos celulares, los núcleos y fibras estructurales clave con un microscopio confocal mientras el embrión se desarrollaba. Se centraron en una ventana temporal en la que las células formaban un anillo alto y ordenado alrededor de la vesícula. En paralelo, examinaron el epidídimo de ratón, un tubo largo y en espiral que ayuda a madurar los espermatozoides, utilizando secciones de tejido finas y marcadores químicos que señalan la síntesis de ADN, las fases del ciclo celular y proteínas estructurales. Este diseño entre especies les permitió probar si la misma danza nuclear ocurre en vertebrados muy distintos.
Las fuerzas que tiran y moldean las células en división
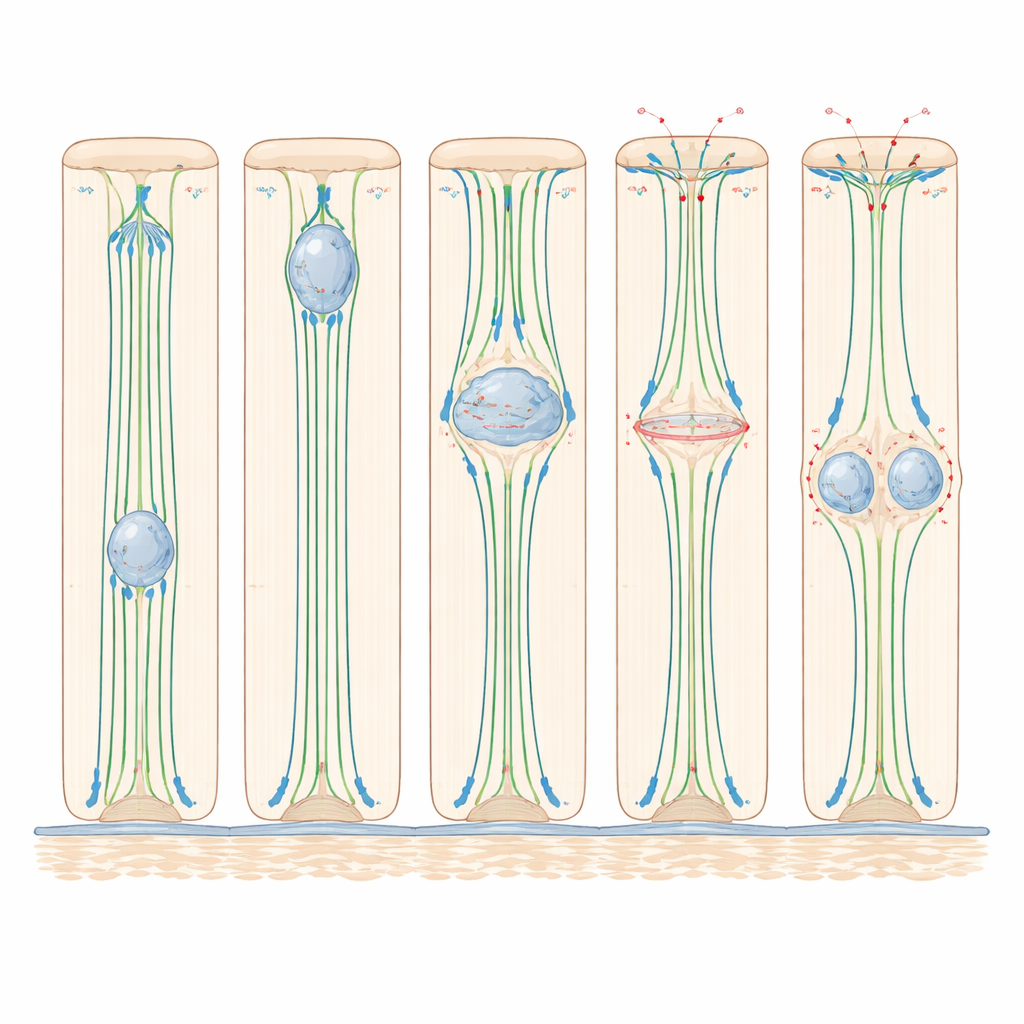
El seguimiento cuidadoso en pez cebra reveló que el núcleo inicia su ascenso solo al final de la fase preparatoria previa a la división y completa el viaje de ida y vuelta en algo más de una hora. Experimentos que interfirieron con distintos “motores” internos mostraron que este movimiento depende en gran medida de los microtúbulos —vías proteicas rígidas que van de la base a la punta— y de la dineína, una proteína motora que se desplaza a lo largo de esas vías hacia la parte superior de la célula. Cuando los microtúbulos se degradaron con fármacos, la migración nuclear se detuvo casi por completo; al bloquear la actividad de la dineína, muchos menos núcleos alcanzaron la cima. En contraste, inhabilitar la miosina II, el principal impulsor de la contracción basada en actina, no ralentizó el ascenso nuclear, lo que revela que en estos epitelios altos pero relativamente simples, el tirón basado en microtúbulos es el motor clave para posicionar el núcleo.
Redondearse sin soltarse
Una vez que el núcleo llega cerca de la superficie interna, la célula experimenta un cambio de forma dramático: se redondea para dividirse. La imagen en vivo y las tinciones proteicas en pez cebra y ratón mostraron que durante este “redondeo” el cuerpo principal de la célula se abomba hacia la luz mientras un delgado tallo de membrana la mantiene anclada a la base. La actina y la miosina II se concentran a lo largo de los costados de la célula, apretándolos como un cordón. Al mismo tiempo, las proteínas que refuerzan las uniones entre células se activan más, ayudando a que la célula en división permanezca firmemente unida a sus vecinas. Cuando se inhibió la miosina II, las células no lograron redondearse correctamente, sus husos a menudo se inclinaron fuera del plano de división plano habitual y uno de los dos núcleos hijas con frecuencia quedó varado cerca de la superficie interna en lugar de reintegrarse en la lámina. Estos resultados muestran que, aunque la miosina II no se necesita para mover el núcleo hacia arriba, sí es crucial para moldear la célula en división y mantener la organización del tejido.
La posición como luz verde para la división
El estudio también descubrió un vínculo estrecho entre la posición del núcleo y si se permite a la célula entrar en la fase final de división. En las células altas del oído del pez cebra, los núcleos rara vez iniciaban mitosis a menos que primero hubieran llegado cerca de la superficie interna. Cuando se perturbaban los microtúbulos, las células en regiones más gruesas del tejido se detenían mayormente antes de dividirse, mientras que las células en regiones más delgadas —donde los núcleos ya estaban cerca de la parte superior— podían continuar. En el epidídimo de ratón, diminutas estructuras llamadas centrosomas permanecían cerca de la superficie interna, y solo cuando el núcleo las alcanzaba los reguladores clave de la división se trasladaban al núcleo y la maquinaria de división se ensamblaba. Esto sugiere que, en estos epitelios, la posición nuclear actúa como un guardián: solo los núcleos que han completado con éxito su viaje ascendente reciben las señales necesarias para comprometerse con la división.
Qué significa esto para tejidos sanos
En conjunto, el trabajo muestra que los epitelios columnares simples en peces y mamíferos comparten una estrategia común: los microtúbulos y la dineína tiran de los núcleos hacia la superficie interna en etapas tardías del ciclo celular; el redondeo impulsado por actomiosina moldea la célula y orienta la división; y mantener un delgado anclaje basal junto con un ángulo de división plano ayuda a que ambas células hijas vuelvan a incorporarse a la capa sin desgarrarla. Al establecer la vesícula ótica del pez cebra y el epidídimo de ratón como modelos complementarios, el estudio ofrece una visión clara y en vivo de cómo las células en división protegen la estructura del tejido —una base para futuras investigaciones sobre cómo estos procesos fallan en la enfermedad y cómo podrían aprovecharse para la reparación tisular.
Cita: Xia, Y., Perder, B., Yao, A.G.C. et al. Zebrafish otic vesicle and mouse epididymis as model systems for studying columnar epithelial cell division. Sci Rep 16, 12995 (2026). https://doi.org/10.1038/s41598-026-42729-z
Palabras clave: división de células epiteliales, migración nuclear, modelo de pez cebra, epidídimo de ratón, imagen celular