Clear Sky Science · de
Zebrafisch-Ohrbläschen und Maus-Epididymis als Modellsysteme zur Untersuchung der Teilung säulenförmiger Epithelzellen
Wie unsere Gewebe sich teilen, ohne auseinanderzufallen
In jedem Moment teilen sich die Zellen, die unsere Organe auskleiden, ersetzen abgenutzte Nachbarn und erhalten gleichzeitig Gewebe wie Darm, Innenohr und Reproduktionstrakt intakt und funktionsfähig. Geht diese Choreografie schief, können Folgeschäden von Fehlbildungen über Unfruchtbarkeit bis hin zu Krebs reichen. Diese Studie stellt zwei starke lebende Modelle vor — ein winziges Fischohr und einen mausspezifischen Fortpflanzungsgang — um in Echtzeit zu beobachten, wie hohe, dicht gepackte Epithelzellen sich teilen, ohne die von ihnen gebildete Barriere zu durchbrechen.
Wenn Zellkerne zur Arbeit pendeln
In vielen Organen sind die Zellen, die dünne Schichten bilden, hoch und säulenförmig, mit einem Ende am Stützgewebe und dem anderen zum flüssigkeitsgefüllten Lumen. Bevor diese Zellen sich teilen, unternehmen ihre Zellkerne einen zeitlich genau abgestimmten „Pendelmarsch“. Sie beginnen nahe der Basis, wandern zur inneren Oberfläche, teilen sich dort und die beiden neuen Kerne reisen dann wieder nach unten. Diese Hin- und Herbewegung, interkinetische nukleare Migration genannt, wurde bisher vor allem im sich entwickelnden Gehirn untersucht. Die Autoren folgerten, dass man, um dieses Verfahren körperweit wirklich zu verstehen, lebende, hochauflösende Einblicke in einfachere, aber dennoch realistische Gewebe benötigt.
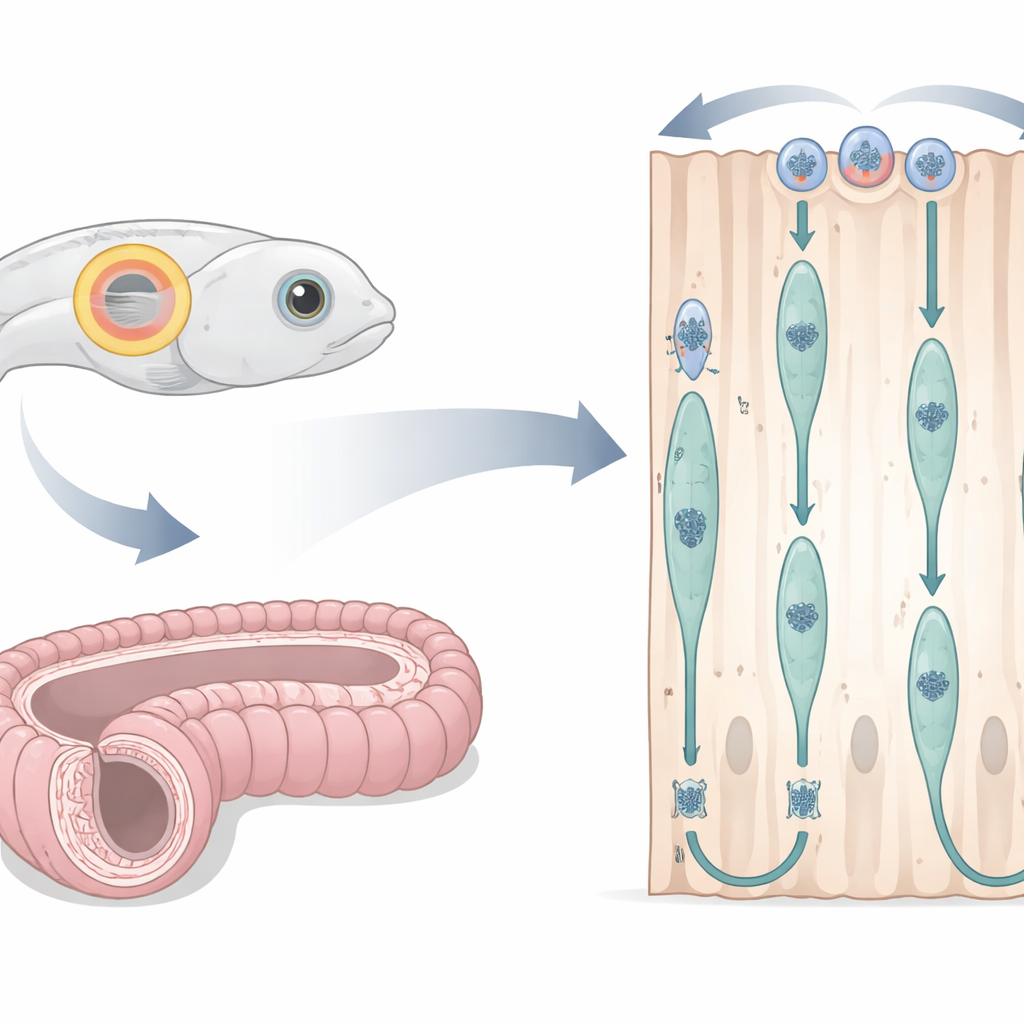
Ein Fischohr und ein Mausgang als lebende Labore
Die Forschenden passten das embryonale Zebrafisch-Otic-Vesikel — einen transparenten, flüssigkeitsgefüllten Sack, der später das Innenohr bildet — als Fenster zur Zellteilung in einfachem säulenförmigem Epithel an. Durch Injektion fluoreszierender Marker in befruchtete Eier konnten sie gleichzeitig Zellumrisse, Zellkerne und wichtige Strukturfasern unter einem Konfokalmikroskop beobachten, während sich das Embryo entwickelte. Sie konzentrierten sich auf ein Zeitfenster, in dem die Zellen einen ordentlichen, hohen Ring um das Bläschen bildeten. Parallel dazu untersuchten sie die Maus-Epididymis, einen langen gewundenen Gang, der zur Spermienreifung beiträgt, mithilfe dünner Gewebeschnitte und chemischer Marker, die DNA‑Synthese, Zellzyklusphasen und Strukturproteine kennzeichnen. Dieses bereichsübergreifende Design erlaubte es ihnen zu prüfen, ob dieselbe Kernbewegung auch bei sehr unterschiedlichen Wirbeltieren abläuft.
Kräfte, die teilende Zellen ziehen und formen
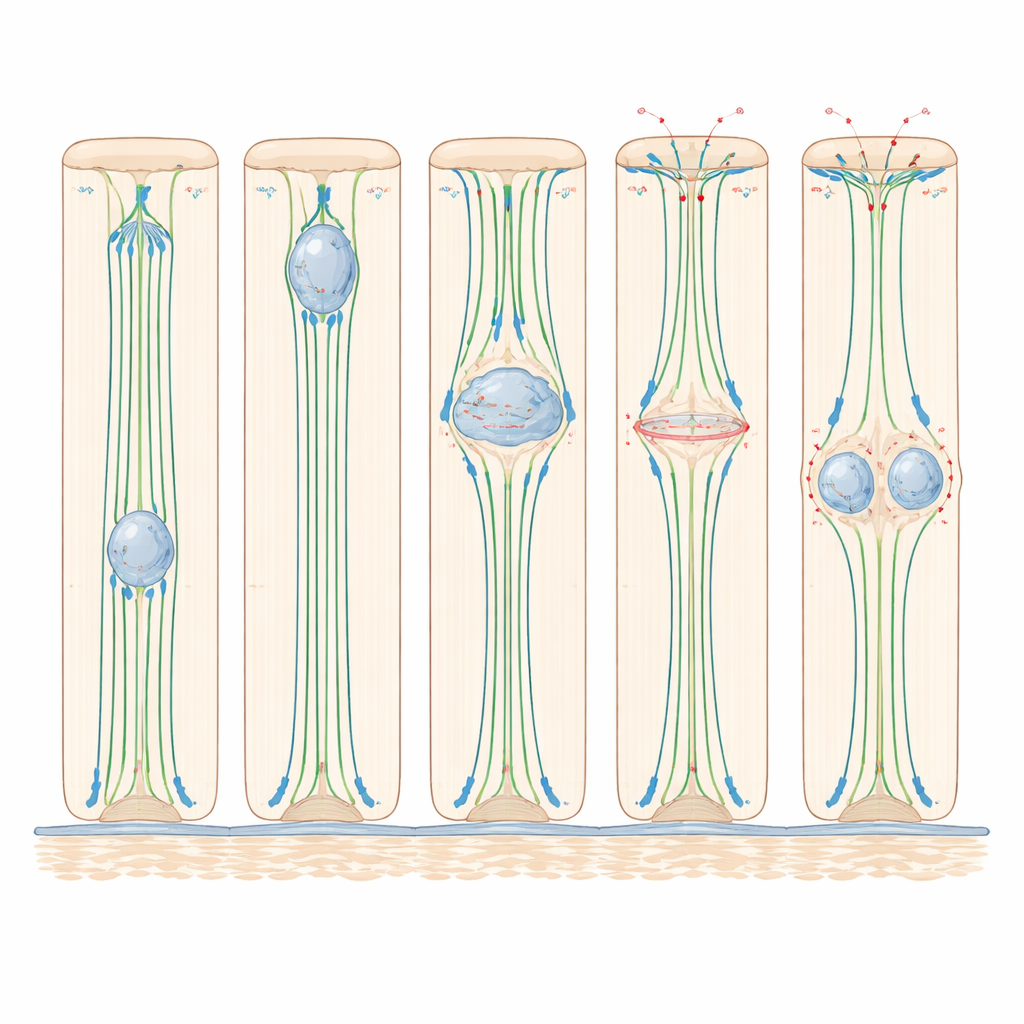
Genaues Tracking im Zebrafisch zeigte, dass der Kern seine Aufwärtsbewegung erst spät in der Vorbereitungsphase vor der Teilung beginnt und die Hin‑ und Rückreise in etwas mehr als einer Stunde abschließt. Experimente, die verschiedene interne „Motoren“ störten, zeigten, dass diese Bewegung stark von Mikrotubuli — starren Proteinbahnen von Basis zu Spitze — und von Dynein abhängt, einem Motorprotein, das entlang dieser Bahnen zur Oberseite der Zelle wandert. Werden Mikrotubuli mit Wirkstoffen abgebaut, stockt die Kernmigration nahezu; wird die Dynein‑Aktivität blockiert, erreichen deutlich weniger Kerne die Spitze. Im Gegensatz dazu verlangsamte die Inaktivierung von Myosin II, dem Haupttreiber aktinbasierter Kontraktion, die Aufwärtsbewegung der Kerne nicht, was zeigt, dass in diesen hohen, aber vergleichsweise einfachen Epithelien das Mikrotubuli‑gestützte Ziehen der entscheidende Motor ist, um den Kern in Position zu bringen.
Abrollen ohne loszulassen
Hat der Kern die Nähe der inneren Oberfläche erreicht, durchläuft die Zelle eine dramatische Formänderung: sie rundet sich zur Teilung auf. Lebendbildaufnahmen und Proteinfärbungen in Zebrafisch und Maus zeigten, dass sich dabei der Zellkörper zum Lumen hin aufbläht, während ein dünner Membranstiel die Verankerung an der Basis erhält. Aktin und Myosin II konzentrieren sich entlang der Zellseiten und ziehen diese wie eine Kordel zusammen. Gleichzeitig werden Proteine, die Zell‑Zell‑Verbindungen stärken, aktiver und helfen der teilenden Zelle, fest an ihren Nachbarn zu haften. Bei Hemmung von Myosin II rundeten Zellen nicht richtig auf, die Teilungsspindeln kippten häufig aus der normalen, flachen Teilungsebene, und einer der beiden Tochterkerne blieb oft in der Nähe der inneren Oberfläche hängen, anstatt wieder in die Zellschicht zu integrieren. Diese Ergebnisse zeigen, dass Myosin II zwar nicht für das Aufwärtsbewegen des Kerns nötig ist, aber entscheidend dafür, die teilende Zelle zu formen und das Gewebe organisiert zu halten.
Position als grünes Licht für die Teilung
Die Studie enthüllte außerdem eine enge Verbindung zwischen der Lage des Kerns und der Frage, ob die Zelle die Erlaubnis erhält, in die finale Teilungsphase einzutreten. In hohen Zebrafisch‑Ohrzellen begann die Mitose selten, wenn die Kerne nicht zuvor die Nähe zur inneren Oberfläche erreicht hatten. Wurden Mikrotubuli gestört, blieben Zellen in dickeren Geweberegionen meist vor der Teilung stecken, während Zellen in dünneren Regionen — wo die Kerne bereits nahe der Spitze lagen — fortfahren konnten. In der Maus‑Epididymis hielten sich winzige Strukturen, die Zentrosomen, in der Nähe der inneren Oberfläche auf, und erst wenn der Kern sie erreichte, bewegten sich zentrale Teilungsregulatoren in den Kern und die Teilungsmaschinerie setzte sich zusammen. Das legt nahe, dass in diesen Epithelien die Kernposition als Torwächter fungiert: Nur Kerne, die ihre Aufwärtsreise erfolgreich abgeschlossen haben, erhalten die Signale, die zur Verpflichtung zur Teilung nötig sind.
Was das für gesunde Gewebe bedeutet
Zusammen genommen zeigt die Arbeit, dass einfache säulenförmige Epithelien bei Fischen und Säugetieren eine gemeinsame Strategie teilen: Mikrotubuli und Dynein ziehen Kerne spät im Zellzyklus zur inneren Oberfläche; aktomyosin‑getriebene Aufrundung formt die Zelle und richtet die Teilung aus; und das Aufrechterhalten einer schlanken basalen Verankerung plus eines flachen Teilungswinkels hilft beiden Tochterzellen, ohne Zerreißen in die Schicht zurückzugleiten. Indem die Studie das Zebrafisch‑Otic‑Vesikel und die Maus‑Epididymis als komplementäre Modelle etabliert, liefert sie eine klare Live‑Ansicht davon, wie teilende Zellen die Gewebestruktur bewahren — eine Grundlage für künftige Forschung dazu, wie diese Prozesse bei Krankheit gestört werden und wie man sie für Gewebeersatz oder Reparatur nutzen könnte.
Zitation: Xia, Y., Perder, B., Yao, A.G.C. et al. Zebrafish otic vesicle and mouse epididymis as model systems for studying columnar epithelial cell division. Sci Rep 16, 12995 (2026). https://doi.org/10.1038/s41598-026-42729-z
Schlüsselwörter: Epithelzellteilung, nukleare Migration, Zebrafischmodell, Maus-Epididymis, Zellbildgebung