Clear Sky Science · en
Zebrafish otic vesicle and mouse epididymis as model systems for studying columnar epithelial cell division
How Our Tissues Divide Without Falling Apart
Every moment, the cells lining our organs divide, replacing worn-out neighbors while keeping tissues like the intestine, inner ear, and reproductive tract intact and functioning. If this choreography goes wrong, the results can include birth defects, infertility, and cancer. This study introduces two powerful living models—a tiny fish ear and a mouse reproductive duct—to watch, in real time, how tall, tightly packed epithelial cells divide without breaking the barrier they form.
Watching Cell Nuclei Take a Daily Commute
In many organs, the cells that form thin sheets are tall and column-shaped, with one end attached to a supporting layer and the other facing a fluid-filled space. Before these cells divide, their nuclei take a carefully timed “commute.” They start near the base, move upward toward the inner surface, divide there, and then the two new nuclei travel back down. This back-and-forth journey, called interkinetic nuclear migration, has been studied mostly in developing brain tissue. The authors reasoned that to really understand how this process works across the body, they needed live, high‑resolution views in simpler but still realistic tissues.
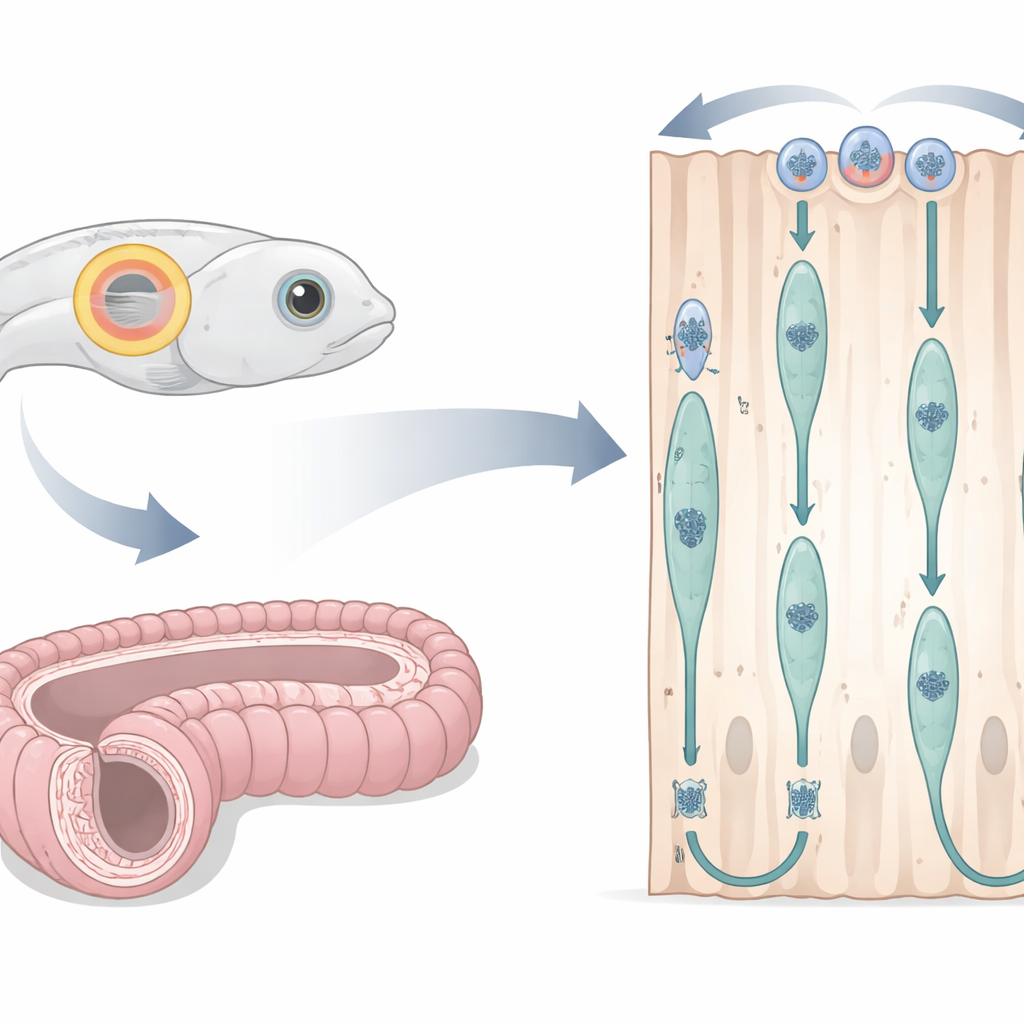
A Fish Ear and a Mouse Duct as Living Laboratories
The researchers adapted the embryonic zebrafish otic vesicle—a transparent fluid-filled sac that will become the inner ear—as a window into cell division in simple columnar epithelium. By injecting fluorescent markers into fertilized eggs, they could simultaneously see cell outlines, nuclei, and key structural fibers under a confocal microscope while the embryo developed. They focused on a time window when cells formed a neat, tall ring around the vesicle. In parallel, they examined the mouse epididymis, a long coiled tube that helps sperm mature, using thin tissue sections and chemical labels that mark DNA synthesis, cell-cycle stages, and structural proteins. This cross-species design allowed them to test whether the same nuclear dance plays out in very different vertebrates.
The Forces that Pull and Shape Dividing Cells
Careful tracking in zebrafish revealed that the nucleus starts its upward journey only late in the preparation phase before division, and completes a round trip in a little over an hour. Experiments that interfered with different internal “motors” showed that this movement depends heavily on microtubules—stiff protein tracks running from base to tip—and on dynein, a motor protein that walks along these tracks toward the top of the cell. When microtubules were broken down with drugs, nuclear migration nearly stopped; when dynein activity was blocked, far fewer nuclei reached the top. In contrast, disabling myosin II, the main driver of actin-based contraction, did not slow the upward nuclear movement, revealing that in these tall but relatively simple epithelia, microtubule-based pulling is the key engine for getting the nucleus into position.
Rounding Up Without Letting Go
Once the nucleus arrives near the inner surface, the cell undergoes a dramatic shape change: it rounds up to divide. Live imaging and protein staining in both zebrafish and mice showed that during this “rounding,” the main cell body balloons toward the lumen while a thin stalk of membrane keeps it anchored to the base. Actin and myosin II concentrate along the sides of the cell, tightening them like a drawstring. At the same time, proteins that strengthen cell–cell junctions become more active, helping the dividing cell stay firmly attached to its neighbors. When myosin II was inhibited, cells failed to round properly, their spindles often tilted away from the normal, flat division plane, and one of the two daughter nuclei frequently became stranded near the inner surface instead of reintegrating into the sheet. These results show that, even though myosin II is not needed to move the nucleus upward, it is crucial for shaping the dividing cell and keeping the tissue organized.
Position as a Green Light for Division
The study also uncovered a tight link between where the nucleus sits and whether the cell is allowed to enter the final division phase. In tall zebrafish ear cells, nuclei rarely started mitosis unless they had first arrived near the inner surface. When microtubules were disrupted, cells in thicker regions of the tissue mostly stalled before division, while cells in thinner regions—where nuclei were already close to the top—could proceed. In the mouse epididymis, tiny structures called centrosomes stayed near the inner surface, and only when the nucleus reached them did key division regulators move into the nucleus and the division machinery assemble. This suggests that, in these epithelia, nuclear position acts as a gatekeeper: only nuclei that have successfully completed their upward journey receive the signals needed to commit to division.
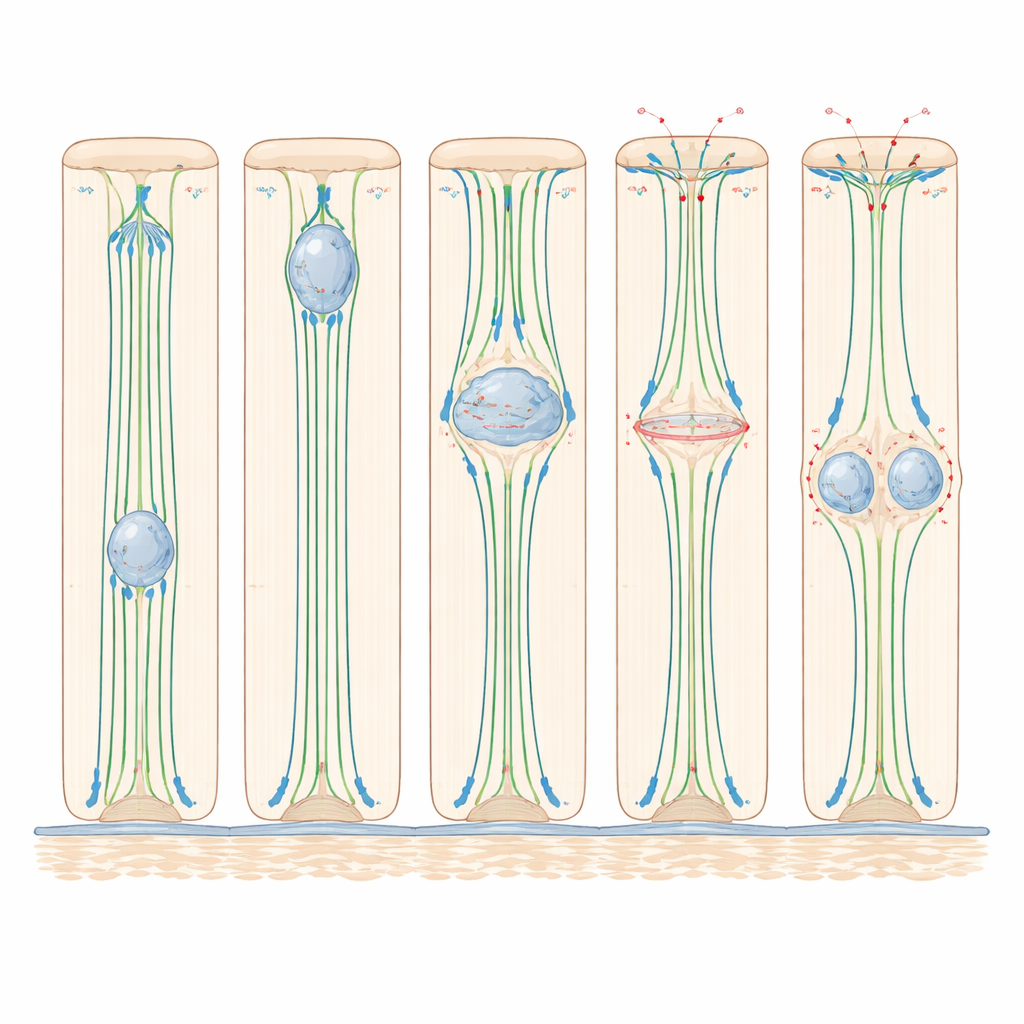
What This Means for Healthy Tissues
Taken together, the work shows that simple columnar epithelia in fish and mammals share a common strategy: microtubules and dynein pull nuclei toward the inner surface late in the cell cycle; actomyosin-driven rounding shapes the cell and orients the division; and maintaining a slim basal attachment plus a flat division angle helps both daughter cells slip back into the layer without tearing it. By establishing the zebrafish otic vesicle and mouse epididymis as complementary models, the study provides a clear, live-action view of how dividing cells safeguard tissue structure—a foundation for future research into how these processes go awry in disease and how they might be harnessed for tissue repair.
Citation: Xia, Y., Perder, B., Yao, A.G.C. et al. Zebrafish otic vesicle and mouse epididymis as model systems for studying columnar epithelial cell division. Sci Rep 16, 12995 (2026). https://doi.org/10.1038/s41598-026-42729-z
Keywords: epithelial cell division, nuclear migration, zebrafish model, mouse epididymis, cell imaging