Clear Sky Science · es
Reconocimiento de ADN superenrollado y control del corte en la topoisomerasa VI
Desenredando el código retorcido de la vida
Dentro de cada célula, el ADN no es una escalera recta y ordenada, sino un hilo largo y flexible que a menudo se tuerce en bobinas y lazos. Estas torsiones son esenciales para empaquetar metros de material genético en espacios microscópicos, pero también generan enredos y nudos que pueden bloquear procesos vitales como la copia y la lectura de los genes. Este estudio revela cómo una máquina molecular, la topoisomerasa VI, reconoce específicamente y corta de forma segura el ADN muy retorcido, ofreciendo una perspectiva sobre cómo las células gestionan el estrés mecánico en su código genético.
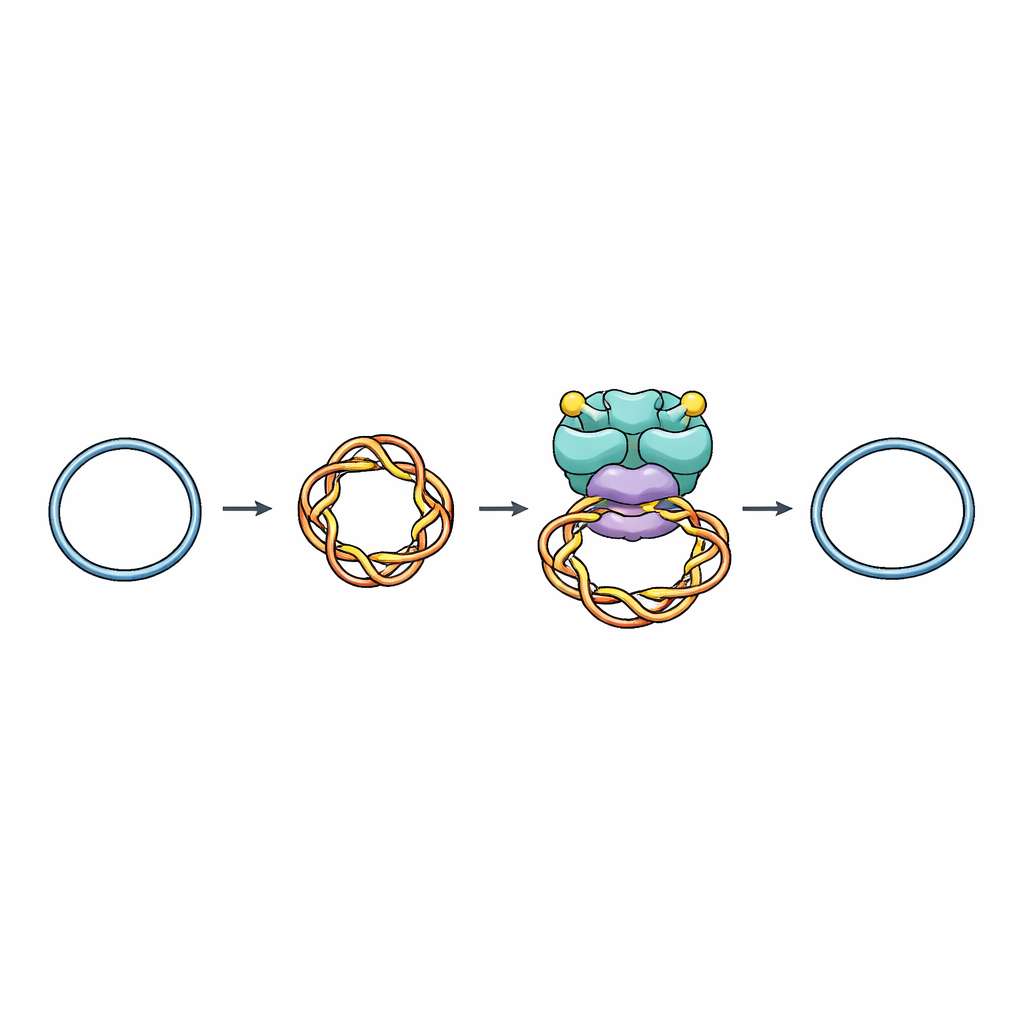
Una herramienta molecular para gestionar los nudos del ADN
Las células dependen de enzimas llamadas topoisomerasas para añadir o quitar torsiones en el ADN, evitando enredos peligrosos durante la replicación, la transcripción y la separación de cromosomas. La topoisomerasa VI (Top6), presente en arqueas y plantas y emparentada con proteínas que inician la meiosis y que defienden a bacterias frente a virus, pertenece a una familia que corta ambas hebras de la doble hélice, hace pasar otro tramo de ADN por la apertura y luego vuelve a sellar la rotura. Hasta ahora, los científicos carecían de una imagen detallada de cómo Top6 reconoce la porción correcta de ADN para cortar y cómo evita realizar roturas accidentales y dañinas.
Capturar al ADN en el acto de torsionarse
Los investigadores utilizaron crio-microscopía electrónica de partícula única, una técnica que obtiene imágenes de moléculas congeladas a resolución casi atómica, para visualizar a Top6 unido a pequeños fragmentos circulares de ADN que se diseñaron para estar superenrollados, como una banda de goma demasiado tensada. Resolvieronn estructuras de Top6 en estados tanto no cortado como cortado mientras la enzima sujetaba estas bobinas. En todos los casos, Top6 abrazaba un tramo de 74 pares de bases de ADN doblado en un lazo cerrado que recuerda a la punta de un plectonema superenrollado, el tipo de doblez agudo que aparece de forma natural en el ADN sobreenrollado. Esto explica por qué Top6 prefiere el ADN superenrollado frente a fragmentos relajados y rectos: su bolsillo de unión está moldeado para encajar un segmento fuertemente curvado.
Cómo la enzima siente y elige el lugar correcto
Una inspección más detallada mostró que Top6 no se limita a agarrar cualquier ADN doblado. Favorece una región particular cuya secuencia subyacente la hace especialmente fácil de deformar. Al analizar los mapas de densidad, el equipo infirió qué pares de bases estaban presentes en el sitio de corte y luego calculó cuán flexibles debían ser los distintos tramos del ADN circular. El sitio de escisión se sitúa en un límite donde un segmento comparativamente rígido se encuentra con otro muy flexible, y los pasos de bases exactos donde se producen los cortes son en sí excepcionalmente plegables. Alrededor de este punto, el ADN se fuerza a adoptar una forma más comprimida y ligeramente no estándar. La enzima parece usar esta combinación de flexibilidad local y doblez agudo como una firma física, que le permite localizar sitios de corte seguros sin “leer” el código genético letra por letra.
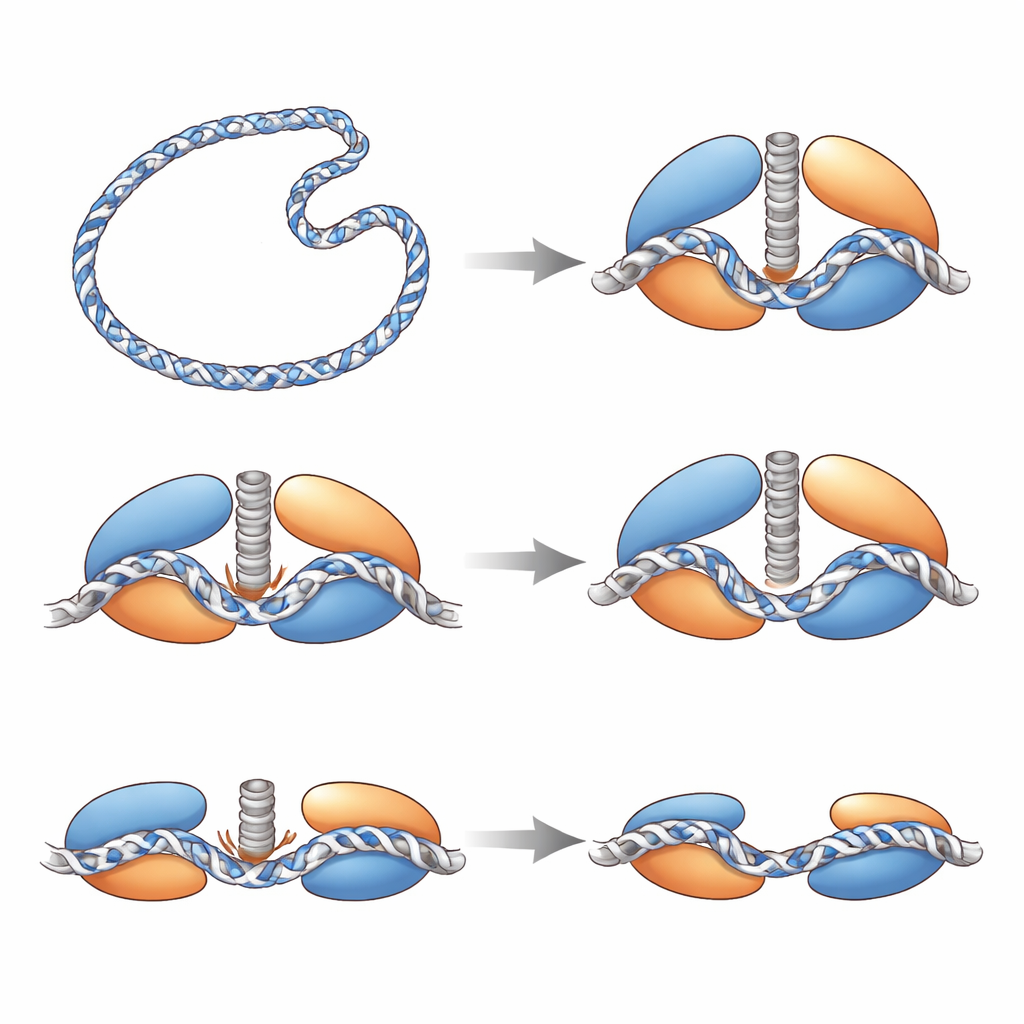
Un pestillo de seguridad y un sensor de tensión
Las estructuras también revelan salvaguardias internas que impiden que Top6 corte el ADN con demasiada facilidad. Dentro de la mitad de la enzima dedicada al procesamiento del ADN, un bucle flexible se ordena solo cuando el ADN está fuertemente doblado en su sitio; actúa como un pestillo que apuntala el sitio activo frente al ADN. Un par de aminoácidos con cargas opuestas forma un “cierro electrostático” que retiene físicamente el residuo de tirosina catalítico para que no alcance la columna vertebral del ADN. Cuando el equipo debilitó este cierre, la enzima produjo muchas más roturas, lo que demuestra que dicho cierre suele suprimir la escisión. En la mitad de Top6 impulsada por ATP, un largo tallo helicoidal que enlaza el movimiento en la región de obtención de energía con la región de corte de ADN se comporta como un sensor de tensión: cuando está intacto y recto, permite una configuración cerrada y lista para el corte; cuando está parcialmente desplegado o desestabilizado por mutación, la enzima tiene dificultades para relajar los superenrollamientos, lo que indica que la flexibilidad controlada de este tallo coordina el uso de la energía con un corte seguro.
El ADN como socio mecánico activo
Al reunir estas observaciones, el estudio dibuja a Top6 y al ADN como socios en una danza mecánica. El superenrollamiento almacena energía en el ADN, impulsando que ciertas secuencias se doblen bruscamente. Top6 reconoce estas formas estresadas, las sujeta, y—solo cuando su pestillo interno, su cierre y su tallo están correctamente posicionados—realiza una rotura controlada de doble hebra antes de volver a sellarla. Este trabajo muestra que las propiedades físicas del ADN, no solo su secuencia de letras, contribuyen a dirigir dónde y cuándo se producen los cortes. Al aclarar cómo Top6 detecta y responde al ADN superenrollado, los hallazgos profundizan nuestra comprensión del mantenimiento de los cromosomas, la recombinación meiótica y algunos sistemas de defensa bacteriana, y subrayan cómo la vida explota la física de las moléculas retorcidas para mantener el genoma compacto e íntegro.
Cita: Richman, D.E., Wendorff, T.J., Rashid, F. et al. Supercoiled DNA recognition and cleavage control in topoisomerase VI. Nat Commun 17, 3092 (2026). https://doi.org/10.1038/s41467-026-69491-0
Palabras clave: superenrollamiento del ADN, topoisomerasa VI, mecánica del ADN, crio-microscopía electrónica, estabilidad del genoma