Clear Sky Science · zh
溶酶体磷酸肌醇脂周转位于RagGTP酶–mTORC1上游并控制肌肉生长
为何微小的细胞枢纽对肌力至关重要
肌肉并非只是因为我们多吃或多锻炼就增长;在每个肌肉细胞内部,微观的控制中心决定是合成新蛋白还是回收旧的成分。本文探讨了其中一个控制中心——溶酶体——如何利用其膜上一类特殊脂质来调节一个强有力的生长开关。通过揭示这一隐藏的检查点,研究者解释了为何一种罕见的儿童肌病会导致严重无力,并展示了针对该通路可以在小鼠中恢复力量。
构建与分解之间的平衡
肌肉细胞不断在两项相对的工作间权衡:合成代谢,构建蛋白并增大肌纤维;以及分解代谢,将组分分解以用于回收。一个重要决策者是位于溶酶体上的蛋白复合物mTORC1。氨基酸等营养物质充足时,mTORC1开启蛋白合成;匮乏时,它被抑制,细胞进入节能模式。本研究关注的是溶酶体外膜的成分如何帮助设定这种平衡,尤以不成熟肌细胞向成熟纤维转变期间为然。 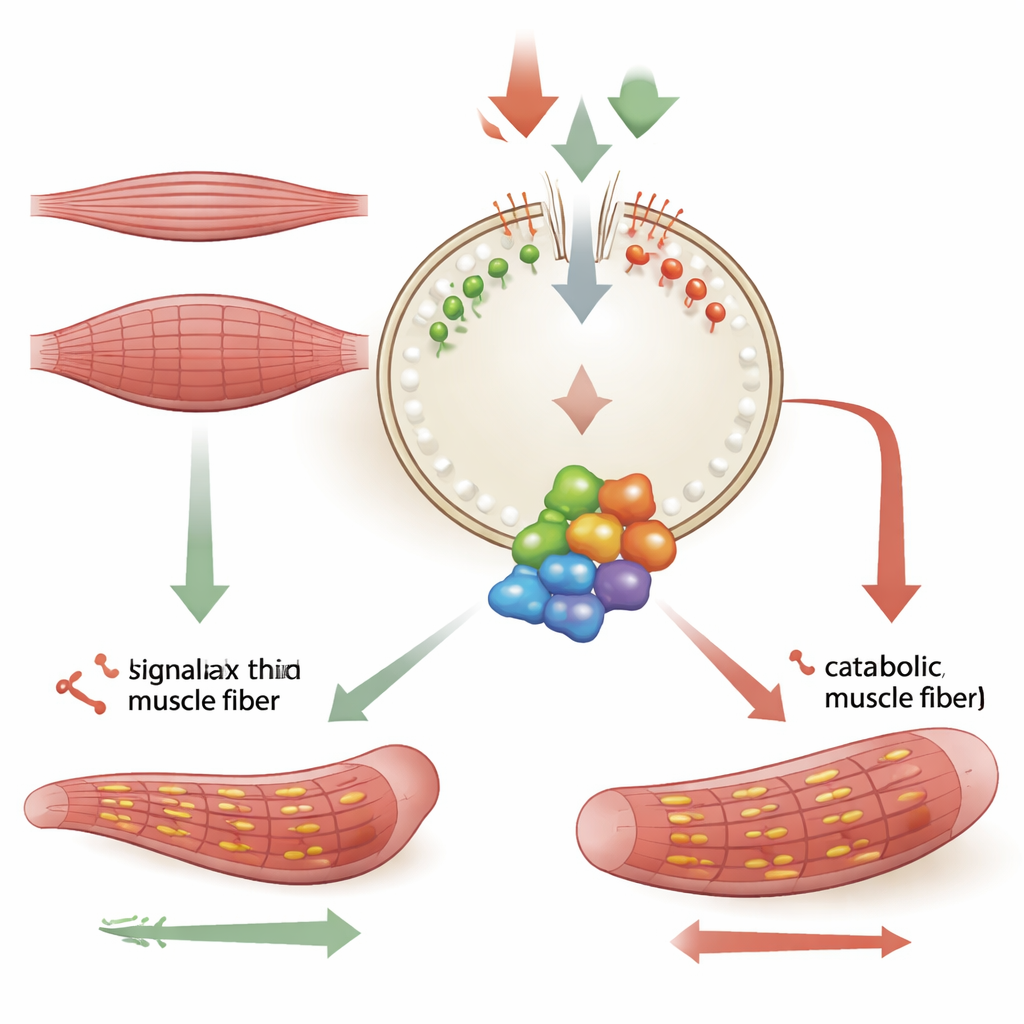
一种罕见肌病揭示了隐藏的开关
团队从一个简单问题出发:哪些与遗传性肌肉萎缩相关的基因会在肌细胞发育过程中改变蛋白合成速率?他们使用小鼠肌肉细胞系,逐一降低36个此类基因的活性,并用荧光氨基酸类似物追踪新蛋白生成。大多数改变影响有限,但失去名为MTM1的基因时,蛋白合成显著增加,尤为突出。MTM1在一种称为X连锁中央核型肌病的严重疾病中发生突变,受影响婴儿会出现严重肌无力。来自患者的细胞和缺乏MTM1的小鼠模型也显示蛋白生成上调,然而形成的肌纤维更薄且融合不良,表明在错误时间“更多”的蛋白合成反而可能损害肌肉生长。
溶酶体上的脂质引导生长通路
MTM1编码一种去除特定膜脂——磷酸肌醇脂——上磷酸基团的酶。作者发现MTM1在溶酶体与内质网接触位点发挥作用。那里,它帮助调控溶酶体表面两种脂质——PI3P和PI(3,5)P2——的水平。当MTM1缺失时,这些脂质在溶酶体上积累。过量的脂质分布吸引并稳定了名为LAMTOR的对接复合物,进而使小型分子开关Rag GTP酶和mTORC1生长机器停留在溶酶体上。结果是mTORC1活动持续偏高,即便在分化某些阶段或轻度应激下细胞本应降低其活性。使用化学或基因方法降低PI3P和PI(3,5)P2的产生,或削弱LAMTOR对溶酶体的结合,能够解除这种过度激活,使肌细胞形成更粗、更健康的纤维。
应激信号与膜接触微调生长
在发育中的肌肉细胞中,内质网在加速蛋白折叠时通常会经历受控水平的应激。在健康细胞中,MTM1通过内质网与溶酶体的接触位点将这种应激转译为对mTORC1的制动,方法是重塑溶酶体脂质。在缺乏MTM1的细胞中,这些接触减少、脂质周转受阻,LAMTOR–Rag复合物在溶酶体上停留更久。研究显示,强制将MTM1直接定位到溶酶体或回送到内质网接触区可以使脂质模式恢复正常,减少mTORC1在溶酶体上的富集,并挽救肌细胞分化。这将MTM1依赖的脂质重塑定位为一种安全阀,防止细胞受压时生长信号失控。 
从细胞生物学到潜在疗法
为检验降低这一过度活跃生长通路是否能帮助病变肌肉,研究者用一种直接抑制mTORC1的药物治疗了缺乏MTM1的小鼠。无论在出现症状前不久开始治疗,还是在肌无力明显后才开始,治疗都改善了体重和肌肉重量,增加了肌纤维尺寸,增强了肌力并延长了寿命,对健康小鼠无害。总体而言,这些发现揭示了一个以溶酶体为中心的代谢检查点:其正常功能依赖专门的膜脂与细胞器接触来维持mTORC1在肌肉生长过程中的平衡;当该机制受损,如在此类肌病中,持续的mTORC1活化反而导致肌肉萎缩。理解并靶向这一电路可能为治疗不仅是这种罕见疾病,还有其他以生长信号失调为特征的病症开辟新途径。
引用: Picot, M., Hifdi, N., Vaucourt, M. et al. Lysosomal phosphoinositide turnover acts upstream of RagGTPase–mTORC1 and controls muscle growth. Nat Metab 8, 624–645 (2026). https://doi.org/10.1038/s42255-026-01484-1
关键词: 溶酶体, mTORC1, 磷酸肌醇脂, 肌肉分化, 肌管样肌病