Clear Sky Science · ru
Обмен фосфоинозитидов на лизосомах действует выше RagGTPase–mTORC1 и контролирует рост мышц
Почему имеют значение крошечные клеточные узлы для силы мышц
Мышцы не растут просто потому, что мы больше едим или тренируемся; внутри каждой мышечной клетки микроскопические центры управления решают, строить ли новые белки или перерабатывать старые. В этой статье рассматривается, как один из таких центров — лизосома — использует особый набор липидов в своей мембране, чтобы настраивать мощный переключатель роста. Раскрывая этот скрытый контрольный пункт, исследователи объясняют, почему редкая детская мышечная болезнь вызывает сильную слабость, и показывают, что воздействие на этот путь может восстановить силу у мышей.
Баланс между сборкой и распадом
Мышечные клетки постоянно балансируют между двумя противоположными задачами: анаболизмом, который наращивает белки и увеличивает волокна, и катаболизмом, который разрушает компоненты для переработки. Главным решающим фактором является белковый комплекс mTORC1, расположенный на лизосомах — небольших мешочках для переработки внутри клетки. Когда питательных веществ, таких как аминокислоты, много, mTORC1 включает синтез белка; при их дефиците он затихает и позволяет клетке перейти в режим экономии. Исследование сосредоточено на том, как состав наружной мембраны лизосомы помогает задавать этот баланс, особенно в процессе превращения незрелых мышечных клеток в зрелые волокна. 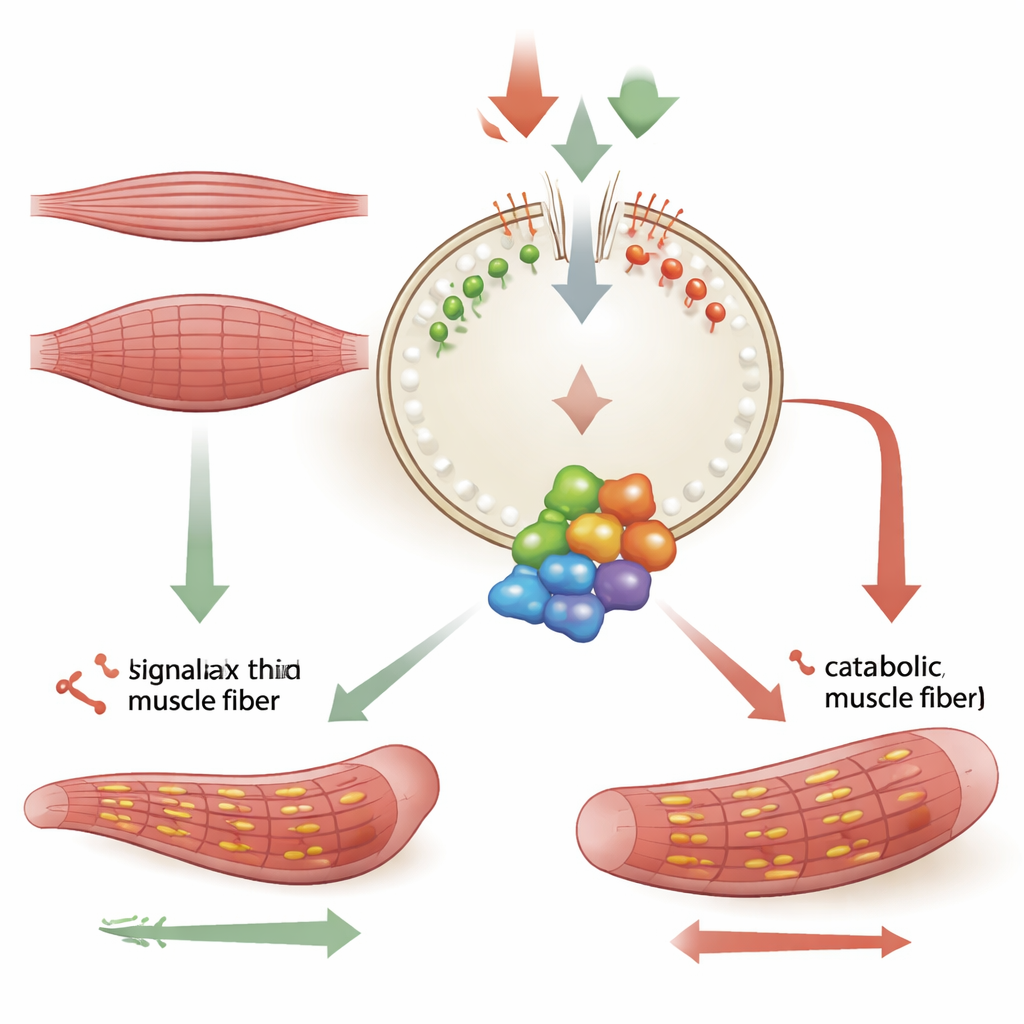
Редкая миопатия выявляет скрытый переключатель
Команда начала с простого вопроса: какие гены, связанные с наследственными расстройствами истощения мышц, изменяют скорость синтеза белка в ходе развития мышечных клеток? Используя мышиную клеточную линию, они поочередно снижали активность 36 таких генов и отслеживали новый синтез белка с помощью флуоресцентных аналогов аминокислот. Большинство изменений давали скромный эффект, но потеря гена MTM1 выделялась — она резко увеличивала синтез белка. MTM1 мутирован при тяжелом состоянии, известном как сцепленная с X центронуклеарная миопатия, при которой у детей развивается глубокая мышечная слабость. Клетки, полученные от пациентов, и мышиная модель с дефицитом MTM1 также демонстрировали повышенное производство белка, но при этом формировали тоньше и плохо сливающиеся мышечные волокна, что указывает на то, что «большее» производство белка в неподходящее время может фактически вредить росту мышц.
Липиды на лизосомах направляют путь роста
MTM1 кодирует фермент, удаляющий фосфатные группы с определенных мембранных липидов, называемых фосфоинозитидами. Авторы обнаружили, что MTM1 действует в контактных зонах, где лизосомы соприкасаются с эндоплазматическим ретикулумом — другой внутренней мембранной сетью. Там он помогает контролировать уровни двух липидов — PI3P и PI(3,5)P2 — на поверхности лизосом. При отсутствии MTM1 эти липиды накапливались на лизосомах. Такой избыток липидов привлекал и стабилизировал докинг-комплекс под названием LAMTOR, который, в свою очередь, удерживал небольшие молекулярные переключатели Rag GTPases и ростовой аппарат mTORC1. В результате активность mTORC1 оставалась стойко высокой, даже когда клетки должны были ее понизить в определенные фазы дифференцировки или при умеренном стрессе. Химические или генетические вмешательства, уменьшающие образование PI3P и PI(3,5)P2 или ослабляющие привязь LAMTOR к лизосоме, снимали это сверхактивирование и позволяли мышечным клеткам формировать более толстые, здоровые волокна.
Сигналы стресса и контакты мембран тонко настраивают рост
Развивающаяся мышца обычно испытывает контролируемый уровень стресса в эндоплазматическом ретикулуме по мере наращивания сворачивания белков. В здоровых клетках MTM1 переводит этот стресс в тормоз для mTORC1, перестраивая липиды лизосом через контактные зоны между двумя органеллами. В клетках с дефицитом MTM1 таких контактов было меньше, оборот липидов замедлялся, и комплексы LAMTOR–Rag дольше задерживались на лизосомах. Исследование показывает, что принудительная локализация MTM1 прямо на лизосомах или возвращение его в зоны контакта с эндоплазматическим ретикулумом нормализовало липидный профиль, уменьшало присутствие mTORC1 на лизосомах и спасало дифференцировку мышечных клеток. Это ставит ремоделирование липидов, зависящее от MTM1, в роль своего рода предохранительного клапана, предотвращающего неконтролируемую сигнальную активацию роста при клеточном давлении. 
От клеточной биологии к потенциальной терапии
Чтобы проверить, поможет ли снижение этой сверхактивной ростовой траектории больным мышцам, исследователи лечили мышей с дефицитом MTM1 препаратом, который напрямую ингибирует mTORC1. Независимо от того, начинали ли лечение незадолго до появления симптомов или после появления слабости, терапия улучшала массу тела и мышц, увеличивала размер волокон, повышала силу и продлевала продолжительность жизни, не причиняя вреда здоровым мышам. В совокупности результаты выявляют лизосомно-центричный метаболический контрольный пункт: работающий, он использует специализированные мембранные липиды и контакты органелл, чтобы удерживать mTORC1 в равновесии в ходе роста мышц; при нарушении, как в этой миопатии, стойкая активация mTORC1 парадоксально приводит к разрушению мышечной ткани. Понимание и таргетирование этой схемы может открыть новые пути лечения не только этой редкой болезни, но и других состояний, связанных с нарушенной регуляцией сигналов роста.
Цитирование: Picot, M., Hifdi, N., Vaucourt, M. et al. Lysosomal phosphoinositide turnover acts upstream of RagGTPase–mTORC1 and controls muscle growth. Nat Metab 8, 624–645 (2026). https://doi.org/10.1038/s42255-026-01484-1
Ключевые слова: лизосома, mTORC1, фосфоинозитиды, дифференцировка мышц, миотубулярная миопатия