Clear Sky Science · zh
用于β(1,3)-葡聚糖顺序催化的构象门控机制
自然的“切糖者”如何推动未来绿色技术
许多微生物通过啃食构成真菌、藻类和植物细胞壁的坚硬天然糖类而生存。这些糖类也是生物燃料和健康产品的有前景原料。本研究揭示了一种新发现的微生物酶如何以高度高效的步进方式抓住并切解一种常见糖——β(1,3)-葡聚糖,打开了改进生物质转化的途径,并深化了我们对肠道微生物处理膳食纤维方式的理解。
近距离观察顽固的天然糖
β(1,3)-葡聚糖是由葡萄糖组成的长链,帮助真菌和藻类维持细胞壁的强度,并使植物能够传递信号和应对胁迫。它们对人类也很有意义:某些形式能作用于免疫系统,另一些则可转化为生物燃料和精细化学品。要利用这些价值,科学家必须了解酶如何将这些链切割成更小的片段。与已经有较多研究的纤维素不同,β(1,3)-葡聚糖长期以来被认为主要由以散发、停顿式方式切割的酶处理。
发现一种沿链“行走”的酶
在本工作中,研究人员从擅长消化复杂植物物质的微生物群中挖掘DNA,鉴定出一个先前未表征的GH158酶家族成员,文中称为GH158(Pro)。他们发现该酶偏爱支链较少的β(1,3)-葡聚糖,也能作用于含有β(1,3)和β(1,4)键的混合链。精细的产物分析显示,它几乎只释放出均一的二糖单元,而不是混合片段。这种产物模式是过程化作用的典型特征:酶一次结合后沿链“行走”,不断切下一个又一个小片段,而不是在每次切割后都脱离。 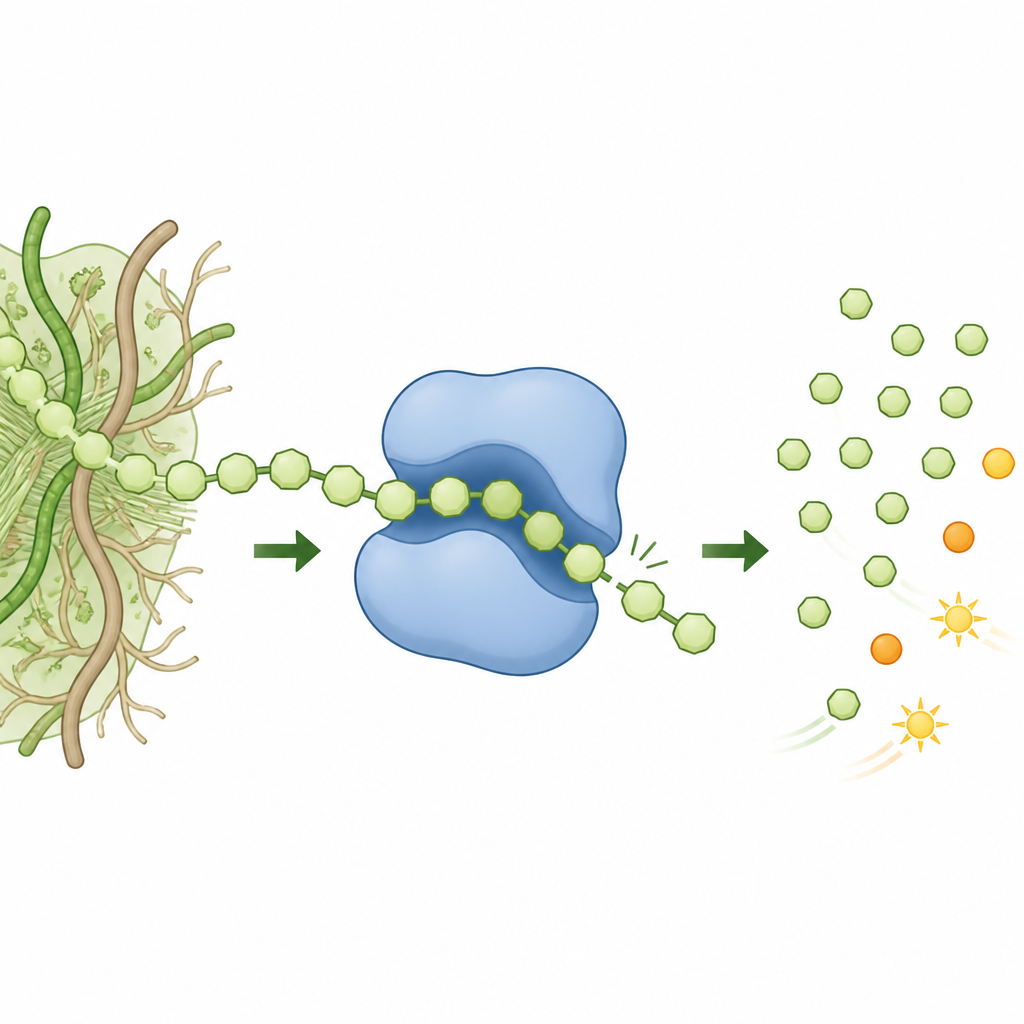
一个移动的隧道抓握并引导糖链
为揭示这种“行走”运动的机制,团队借助先进的X射线方法解析了酶在有无结合糖片段情况下的十九个超高分辨率三维结构。这些快照显示,当底物结合时,酶的一部分折叠覆盖形成一段短隧道,将糖链包裹起来。两个关键区域——一个来自主结构域,一个来自连接的Ig样结构域——互相靠拢,并通过正负电荷氨基酸之间的盐桥被锁定。在隧道内,芳香侧链与弯曲的糖骨架发生堆叠,选择性识别特定位点的β(1,3)连接,同时在更外侧容许β(1,3)或β(1,4)。破坏维持隧道的残基会削弱活性并改变产物分布,使酶的行为更像常见的非过程性切割酶。
开合如何驱动步进式切割
计算机模拟表明该隧道并非刚性。在游离酶中,隧道倾向于打开,而结合糖则稳定封闭构象。切割发生后,对长糖片段的模拟显示,新释放的二糖产物会迅速从隧道的正端退出。随后,盐桥断裂使隧道打开,让剩余链前移一到两个糖单元。当隧道重新闭合时,其他残基重新定位以将链固定到新的、准备切割的构型。量子力学计算进一步表明,被切割的糖在反应过程中遵循一种“循环”的形变,始终以相同的放松构象开始和结束——这种行为此前主要在从链端修剪的酶中观察到。 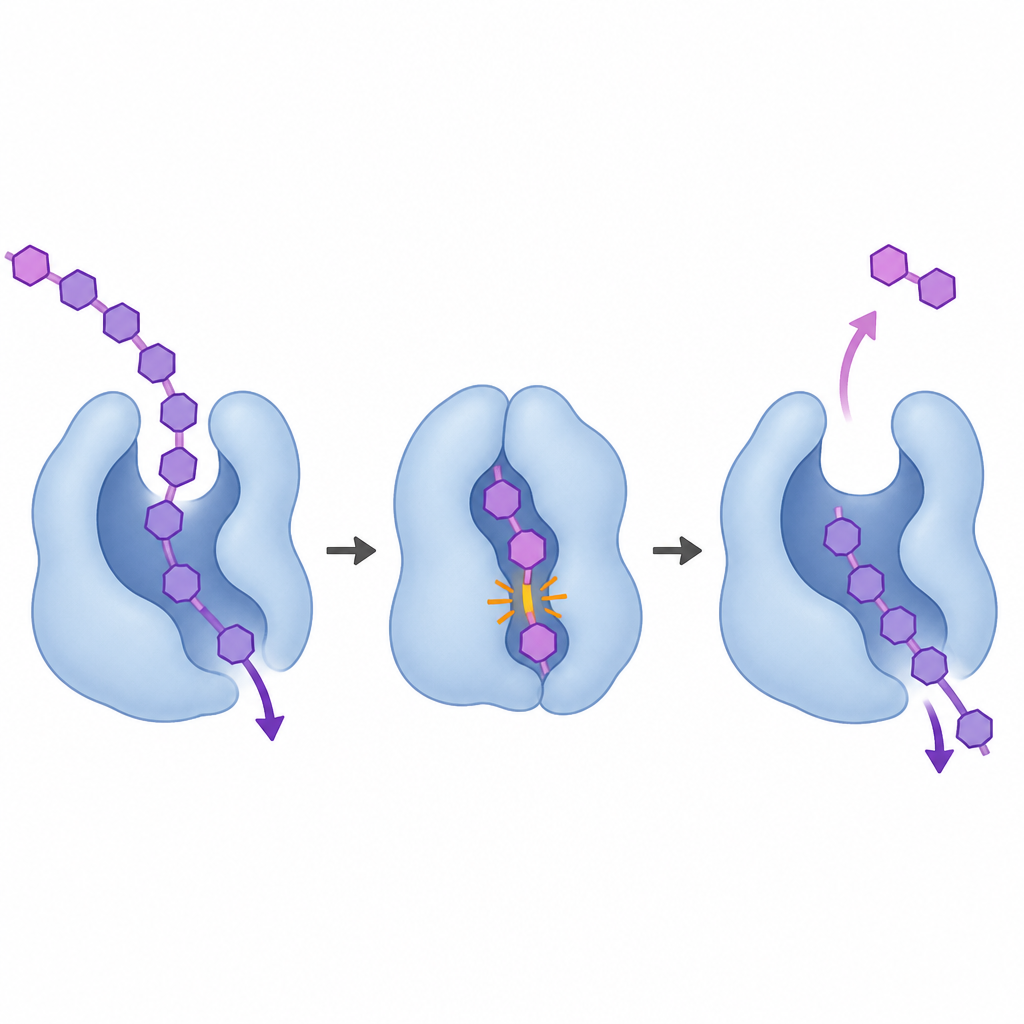
为何这个可动门很重要
这项研究表明,GH158(Pro)中的动态“门控”隧道使一种罕见的内切-过程性策略成为可能,用以分解β(1,3)-葡聚糖。通过在开与闭之间循环,酶既能牢牢抓住链以实现高效切割,又能在不放手的情况下将其前移。作者还发现形成隧道的关键残基在许多相关酶中是保守的,表明这种策略可能很普遍。对普通读者来说,其要点是:自然利用了一个巧妙的可动门,把顽固的细胞壁糖切成整齐统一的小块——这一知识可用于设计更好的酶,以推动可持续燃料、绿色化学,甚至开发与我们微生物群相互作用的定制膳食纤维。
引用: Gimenis, G.H.B., Spadeto, J.P.M., Colombari, F.M. et al. Conformational gating mechanism for processive catalysis of β(1,3)-glucans. Nat Commun 17, 4527 (2026). https://doi.org/10.1038/s41467-026-71224-2
关键词: β-葡聚糖, 过程性酶, 生物质降解, 糖苷水解酶, 生物燃料酶