Clear Sky Science · pl
Mechanizm bramkowania konformacyjnego w procesywnej katalizie β(1,3)-glukanów
Jak przyrodnicze „noże do cukru” napędzają przyszłe zielone technologie
Wiele mikroorganizmów przetrwa, skubiąc twarde naturalne cukry tworzące ściany komórkowe grzybów, glonów i roślin. Te cukry są także obiecującymi surowcami do produkcji biopaliw i produktów zdrowotnych. Badanie to wykazuje, jak nowo odkryty enzym mikrobiologiczny chwyta i rozcina powszechny cukier β(1,3)-glukan w wysoce wydajny, etapowy sposób, otwierając możliwości lepszej konwersji biomasy i pogłębiając zrozumienie, jak nasze własne mikroby jelitowe przetwarzają błonnik.
Bliższe spojrzenie na oporne naturalne cukry
β(1,3)-glukany to długie łańcuchy glukozy, które wzmacniają ściany komórkowe grzybów i glonów oraz pozwalają roślinom przekazywać sygnały i reagować na stres. Interesują też ludzi, ponieważ niektóre formy oddziałują z układem odpornościowym, a inne można przekształcić w biopaliwa i substancje chemiczne wysokiej klasy. Aby wykorzystać te korzyści, naukowcy muszą zrozumieć, jak enzymy rozdrabniają te łańcuchy na mniejsze fragmenty. W odróżnieniu od lepiej poznanej celulozy, której rozkład jest już szeroko badany, β(1,3)-glukany uważano dotąd za rozkładane głównie przez enzymy tnące w sposób rozproszony, przerywany.
Odkrycie enzymu, który „chodzi” wzdłuż łańcucha
W tej pracy badacze przeszukali DNA społeczności mikroorganizmów znanej z doskonałego trawienia złożonego materiału roślinnego i zidentyfikowali wcześniej nieopisany członek rodziny enzymów GH158, nazwany tutaj GH158(Pro). Stwierdzili, że enzym ten preferuje β(1,3)-glukany o niewielkiej liczbie rozgałęzień i może także działać na niektóre mieszaniny łańcuchów zawierających zarówno wiązania β(1,3), jak i β(1,4). Dokładna analiza produktów wykazała, że uwalnia niemal wyłącznie drobne dwucukrowe jednostki, zamiast mieszaniny fragmentów. Taki wzór to cecha procesu procesywnego: enzym zakotwicza się raz, a potem „kroczy” wzdłuż łańcucha, odcinając kolejno małe kawałki, zamiast odrywać się po każdym cięciu. 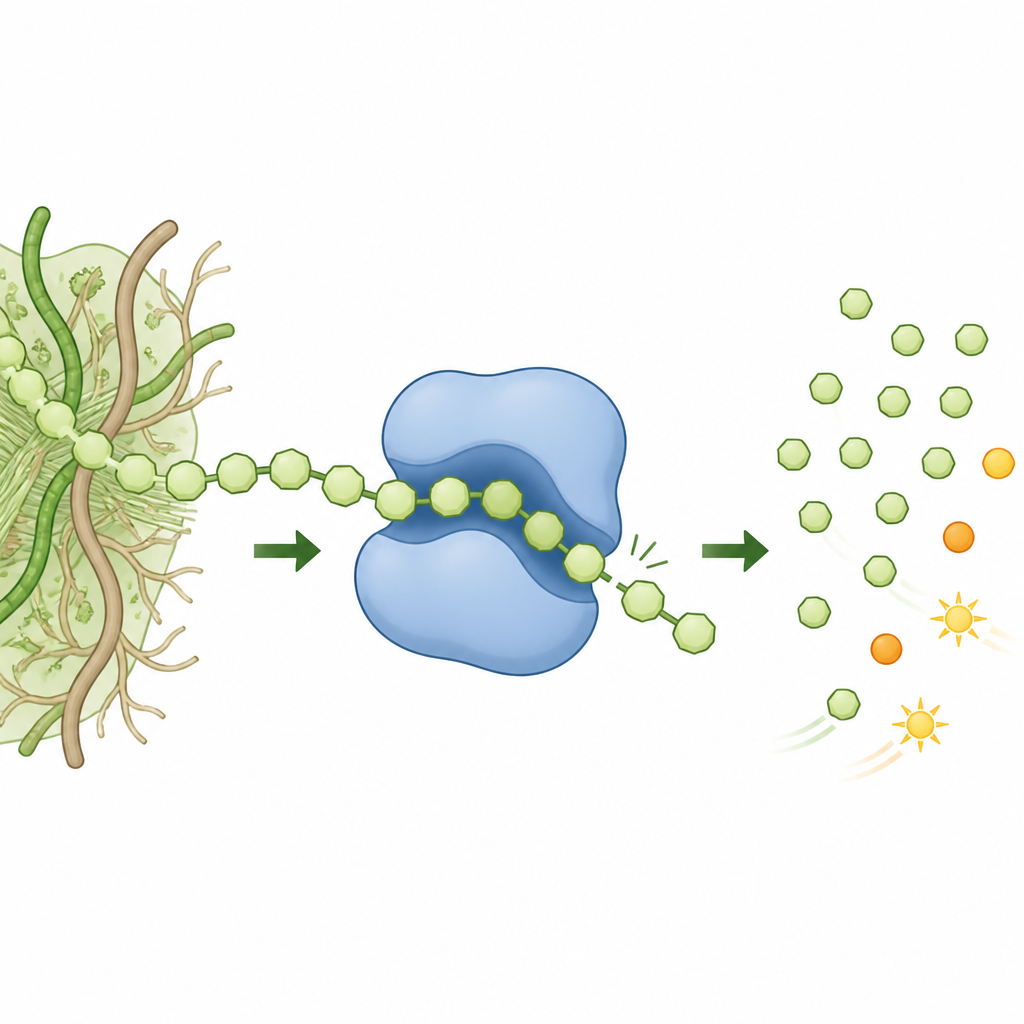
Ruchomy tunel, który chwyta i prowadzi cukier
Aby odkryć, jak działa to „chodzenie”, zespół rozwiązał dziewiętnaście bardzo szczegółowych struktur 3D enzymu z przyłączonymi i bez przyłączonych fragmentów cukrowych, wykorzystując zaawansowane metody rentgenowskie. Te migawki pokazały, że po związaniu substratu część enzymu składa się nad łańcuchem, tworząc krótki tunel otaczający łańcuch cukrowy. Dwa kluczowe obszary — jeden z korpusu enzymu i drugi z dołączonej domeny przypominającej Ig — zbliżają się do siebie i są blokowane przez mostek solny między dodatnio oraz ujemnie naładowanym aminokwasem. Wewnątrz tunelu aromatyczne łańcuchy boczne układają się w stos w stosunku do zakrzywionego szkielety cukrowego, wybierając wiązania β(1,3) w określonych pozycjach, jednocześnie tolerując dalej położone wiązania β(1,3) lub β(1,4). Mutacje reszt utrzymujących tunel osłabiły aktywność i zmieniły skład produktów, powodując, że enzym zachowywał się bardziej jak konwencjonalny, nieprocesywny tnący enzym.
Jak otwieranie i zamykanie napędza etapowe cięcie
Symulacje komputerowe wykazały, że tunel nie jest sztywny. W stanie wolnym tunel ma skłonność do otwierania się, natomiast związanie cukru stabilizuje formę zamkniętą. Po wykonanym cięciu symulacje długich fragmentów cukrowych pokazały, że świeżo uwolniony dwucukrowy produkt szybko opuszcza dodatni koniec tunelu. Następnie rozerwanie mostka solnego pozwala tunelowi się otworzyć, co umożliwia przesunięcie pozostałego łańcucha do przodu o jedną lub dwie jednostki cukrowe. Gdy tunel ponownie się zamyka, inne reszty zmieniają pozycję, aby utrzymać łańcuch w nowej, gotowej do cięcia konfiguracji. Obliczenia kwantowo‑mechaniczne wykazały ponadto, że cięty cukier przechodzi podczas reakcji przez „cykliczną” zmianę kształtu, zaczynając i kończąc w tej samej zrelaksowanej formie — zachowanie wcześniej obserwowane głównie w enzymach przycinających łańcuchy od końców.
Dlaczego ta ruchoma bramka ma znaczenie
Badanie pokazuje, że dynamiczny „bramkowy” tunel w GH158(Pro) umożliwia rzadką endo‑procesywną strategię rozkładu β(1,3)-glukanów. Przełączając się między stanami otwartym i zamkniętym, enzym może zarówno mocno chwycić łańcuch dla wydajnego cięcia, jak i przesunąć go do przodu bez zwalniania uchwytu. Autorzy wykazują też, że kluczowe reszty tworzące tunel są zachowane w wielu powiązanych enzymach, co sugeruje, że ta strategia jest powszechna. Dla czytelnika popularnonaukowego główny wniosek jest taki, że natura używa sprytnej ruchomej bramki, aby przekształcać oporne cukry ścian komórkowych w równe, jednolite kawałki — wiedza ta może zostać wykorzystana do projektowania lepszych enzymów dla zrównoważonych paliw, zielonej chemii, a być może także do tworzenia celowanych włókien pokarmowych oddziałujących z naszą mikrobiotą w określony sposób.
Cytowanie: Gimenis, G.H.B., Spadeto, J.P.M., Colombari, F.M. et al. Conformational gating mechanism for processive catalysis of β(1,3)-glucans. Nat Commun 17, 4527 (2026). https://doi.org/10.1038/s41467-026-71224-2
Słowa kluczowe: beta glukan, enzym procesywny, degradacja biomasy, glikozydaza, enzymy do biopaliw