Clear Sky Science · ru
Нарушенная загрузка когезина нарушает дифференцировку поджелудочной железы посредством перестройки хроматина, управляемой Polycomb, и коллапса петель
Как клетки складывают свою ДНК, чтобы стать клетками поджелудочной железы
В каждой клетке два метра ДНК нужно сложить и снова развернуть так, чтобы нужные гены включались в нужное время. В этой работе исследуется, как этот процесс укладки помогает человеческим стволовым клеткам становиться инсулин-продуцирующими клетками поджелудочной железы — и что происходит, когда отсутствует ключевой «помощник» в укладке. Поскольку ошибки в этой системе связаны с нарушениями развития и, возможно, с диабетом, понимание этой невидимой хореографии ДНК имеет широкие последствия для здоровья.
Машина клетки по сворачиванию ДНК в петли
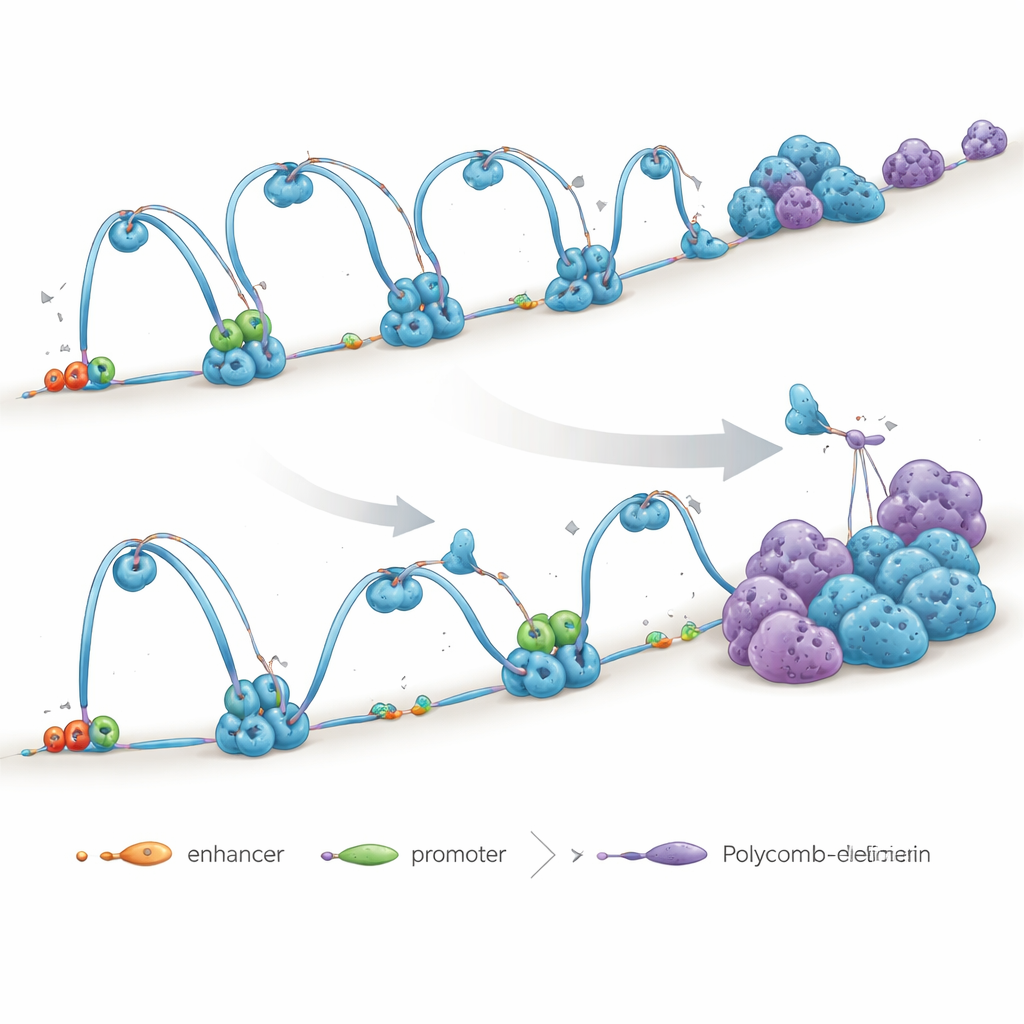
Главное внимание в исследовании уделено белковому помощнику под названием NIPBL, который загружает на ДНК кольцевой комплекс, известный как когезин. Когезин действует как скользящая скоба, которая подтягивает удалённые участки генома в петли, сближая регуляторы генов (усилители) с контролируемыми ими генами (промоторами). Другой белок, CTCF, часто отмечает границы этих петель, помогая вырезать геном на изолированные «районы». Авторы показывают, что NIPBL важен не только для установки когезина на ДНК, но и для поддержания непрерывного процесса «выдавливания петель», который сохраняет целостность этих районов.
Что происходит, когда загрузчик не справляется
Чтобы выяснить роль NIPBL, исследователи уменьшили его уровень в человеческих эмбриональных стволовых клетках. Удивительно, но общее количество когезина во многих пограничных участках осталось высоким, однако дальнодействующие петли, соединяющие удалённые сегменты ДНК, ослабли или исчезли. Контакты усилитель–промотор, необходимые для включения генов, пострадали особенно сильно. Даже там, где когезин сохранился, петли стали короче и менее эффективны, а некоторые большие петли, казалось, коллапсировали в более мелкие локальные структуры. Это показывает, что простого наличия когезина недостаточно — его корректная загрузка и движение по ДНК, обеспечиваемые NIPBL, и есть то, что поддерживает 3D-проводку генома.
Кластеры Polycomb берут верх
ДНК также организуется комплексами репрессии, известными как Polycomb, которые собирают группы генов в подавленные компартменты. Когда петли, управляемые когезином, ослабли после утраты NIPBL, авторы наблюдали противоположную динамику у доменов Polycomb: они взаимодействовали сильнее и формировали более яркие, плотные фокусы в ядре. Дальнодействующие контакты между регионами, богатыми Polycomb, усилились, хотя общее количество Polycomb на ДНК изменилось незначительно. Химические обработки, нарушающие каплеобразные конденсаты или мешающие Polycomb «считывать» специфические химические метки, снижали эти контакты, что указывает на то, что Polycomb использует механизм кластеризации, похожий на фазовое разделение, который становится более доминирующим при нарушении выдавливания петель. Другими словами, когда организация на основе петель слабеет, на передний план выходит компартментная репрессия.
Нарушенные пути к панкреатической идентичности
Команда затем проследила за стволовыми клетками по мере их направления через несколько стадий к образованию панкреатических островковых органоидов, включая бета‑подобные клетки, продуцирующие инсулин. Обычно этот процесс включает открытие специфичных для стадии регуляторных участков ДНК и построение новых петель, соединяющих эти области с панкреатическими генами. Когда NIPBL был снижен в ходе дифференцировки, тысячи генов не достигли нормального уровня активности, и многие участки ДНК, которые должны были стать доступными, оставались закрытыми. Новообразованные петли, характерные для панкреатических прогениторов, стали реже, короче и слабее, особенно те, которые окружали усилители и ключевые гены развития. Ингибирование Polycomb частично восстанавливало активность генов, но не могло восстановить отсутствующие петли, что подчёркивает: образование петель, зависящее от NIPBL, — отдельный и незаменимый уровень управления.
Супер‑переключатели и коллапсированные цепи
Авторы также изучали супер‑усилители — большие кластеры регуляторных элементов, выступающие как «супер‑переключатели» для генов, определяющих идентичность клетки. В нормальных клетках эти области связаны между собой и с целевыми генами посредством петель, опосредованных когезином. После потери NIPBL метки когезина на многих таких супер‑усилителях уменьшились, и петли, соединяющие их, ослабли, в то время как некоторые дальние контакты между разными регионами супер‑усилителей увеличились в более неорганизованной манере. Это говорит о том, что при разрыве обычных цепей петель геном может компенсировать, формируя более широкие, менее точные соединения, что потенциально размывает чёткий механизм включения/выключения, необходимый для генов развития.
Почему это важно для развития и болезней
В целом исследование рисует образ NIPBL как главного организатора 3D‑генома во время решения судьбы клетки. Загружая и мобилизуя когезин, он формирует и поддерживает петли, которые обеспечивают общение усилителей и промоторов, одновременно противодействуя кластеризации репрессированных Polycomb‑регионов. Когда функция NIPBL нарушена, петли усилитель–промотор коллапсируют, компартменты Polycomb уплотняются, и аккуратно скоординированные генетические программы, необходимые для дифференцировки поджелудочной железы, дают сбой. Эта механистическая картина помогает объяснить, как мутации в генах, связанных с когезином, могут вызывать сложные синдромы развития, и даёт подсказки о том, как тонкие нарушения в укладке генома могут способствовать таким заболеваниям, как диабет.
Цитирование: Yu, L., Liu, Y., Zhang, J. et al. Impaired cohesin loading disrupts pancreatic differentiation by Polycomb-driven chromatin rewiring and loop collapse. Commun Biol 9, 590 (2026). https://doi.org/10.1038/s42003-026-09838-x
Ключевые слова: 3D-организация генома, когезин и NIPBL, домены Polycomb, дифференцировка поджелудочной железы, петли усилитель–промотор