Clear Sky Science · pl
Upośledzone ładowanie cohesin zakłóca różnicowanie trzustki przez przegrupowanie chromatyny kierowane przez Polycomb i załamanie pętli
Jak komórki składują swoją DNA, by stać się komórkami trzustki
W każdej komórce dwa metry DNA muszą być składane i ponownie składane, aby właściwe geny włączały się we właściwym czasie. Artykuł bada, jak ten proces składania pomaga ludzkim komórkom macierzystym stać się komórkami trzustki produkującymi insulinę — i co się dzieje, gdy braknie kluczowego pomocnika w tym procesie. Ponieważ błędy w tym systemie wiążą się z zaburzeniami rozwojowymi, a być może z cukrzycą, zrozumienie tej niewidocznej choreografii DNA ma szerokie konsekwencje dla zdrowia.
Maszyna komórkowa do tworzenia pętli DNA
Głównym punktem badania jest białkowy pomocnik o nazwie NIPBL, który ładuje na DNA pierścieniowy kompleks znany jako cohesin. Cohesin działa jak ślizgająca się obejma, która przyciąga odległe fragmenty genomu razem tworząc pętle, zbliżając w ten sposób przełączniki genów (enhancery) do genów, które kontrolują (promotory). Inne białko, CTCF, często wyznacza granice tych pętli, pomagając dzielić genom na izolowane sąsiedztwa. Autorzy pokazują, że NIPBL jest kluczowy nie tylko do umieszczania cohesin na DNA, ale także do napędzania ciągłej „ekstruzji pętli”, która utrzymuje te sąsiedztwa w całości.
Co się dzieje, gdy „ładowacz” zawodzi
Aby zobaczyć, co naprawdę robi NIPBL, badacze zmniejszyli jego poziomy w ludzkich embrionalnych komórkach macierzystych. Ku zaskoczeniu, ogólna ilość cohesin w wielu miejscach granicznych pozostała wysoka, jednak długodystansowe pętle łączące odległe segmenty DNA osłabły lub zniknęły. Kontakty enhancer–promotor, kluczowe dla włączania genów, zostały szczególnie dotknięte. Nawet tam, gdzie cohesin pozostał obecny, pętle stały się krótsze i mniej efektywne, a niektóre duże pętle wydawały się zapadać w mniejsze, lokalne struktury. To pokazuje, że obecność cohesin sama w sobie nie wystarcza — to jego prawidłowe załadowanie i przesuwanie się po DNA, napędzane przez NIPBL, utrzymuje trójwymiarowe okablowanie genomu.
Przejęcie przez klastry Polycomb
DNA jest też organizowane przez kompleksy wyciszające zwane Polycomb, które skupiają grupy genów w zrepresjonowane przegrody. Gdy pętle napędzane przez cohesin osłabły po utracie NIPBL, autorzy zaobserwowali odwrotne zachowanie domen Polycomb: zaczęły one silniej wchodzić w interakcje i tworzyć jaśniejsze, gęstsze ogniska w jądrze. Długodystansowe kontakty między regionami bogatymi w Polycomb wzrosły, mimo że całkowita ilość Polycomb na DNA zmieniła się niewiele. Zabiegi chemiczne, które zaburzają kondensaty przypominające krople lub interferują z zdolnością Polycomb do „czytania” specyficznych oznaczeń chemicznych, zmniejszały te kontakty, co wskazuje, że Polycomb wykorzystuje mechanizm klastrowania przypominający separację fazową, który staje się bardziej dominujący, gdy ekstruzja pętli jest upośledzona. Innymi słowy, gdy organizacja oparta na pętlach zanika, organizacja oparta na kompartmentach wyciszających wkracza do akcji.
Zakłócone drogi prowadzące do tożsamości trzustkowej
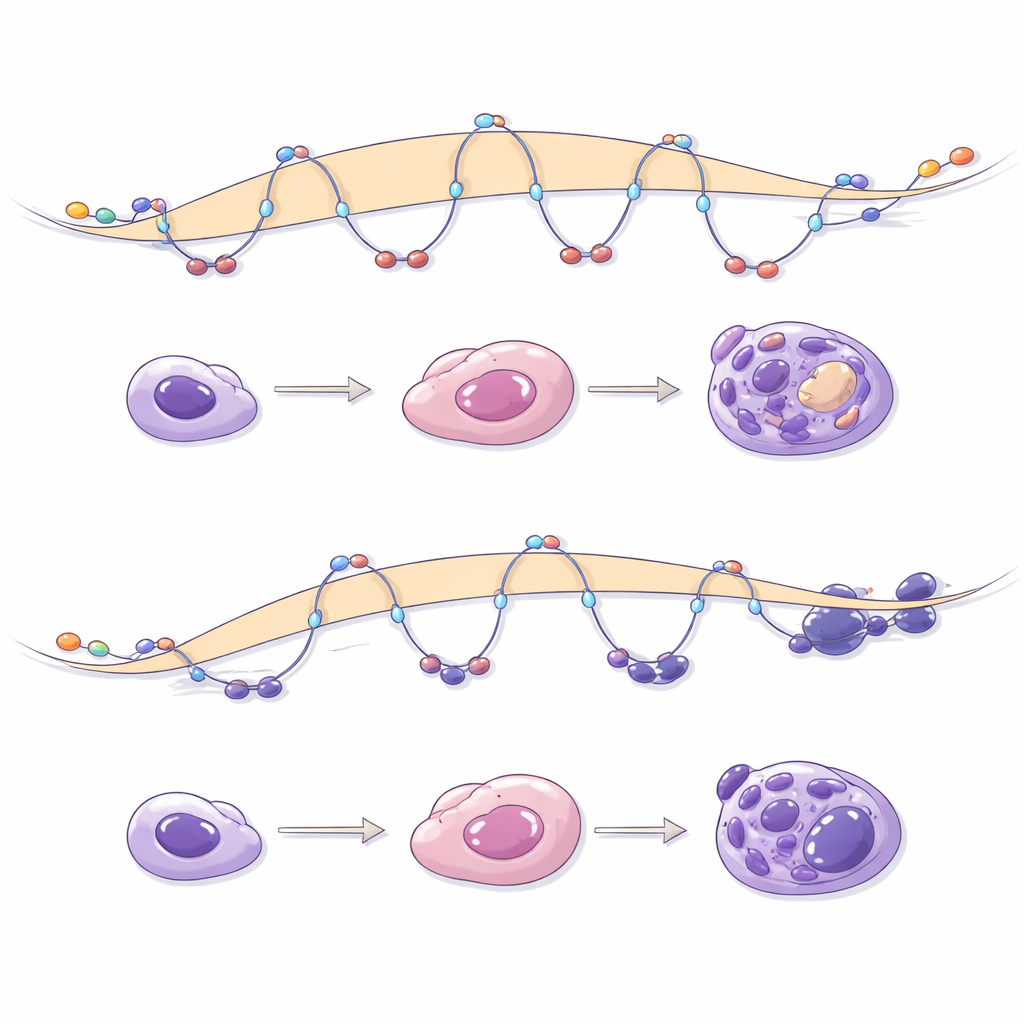
Zespół następnie śledził komórki macierzyste prowokowane przez kilka etapów ku tworzeniu organoidów wysp trzustkowych, w tym komórek przypominających beta produkujących insulinę. Zwykle ten proces obejmuje otwieranie się regulatorych sekwencji DNA specyficznych dla danego etapu i budowanie nowych pętli łączących te regiony z genami trzustkowymi. Gdy NIPBL był zredukowany podczas różnicowania, tysiące genów nie osiągnęło właściwych poziomów aktywności, a wiele miejsc w DNA, które powinny stać się dostępne, pozostało zamkniętych. Nowo powstające pętle typowe dla progenitorów trzustkowych było mniej, były krótsze i słabsze, szczególnie te obejmujące enhancery i kluczowe geny rozwojowe. Hamowanie Polycomb mogło częściowo przywrócić aktywność genów, ale nie potrafiło odbudować brakujących pętli, podkreślając, że tworzenie pętli zależnych od NIPBL jest odrębną i nie do zastąpienia warstwą kontroli.
Super-wyłączniki i zapadnięte obwody
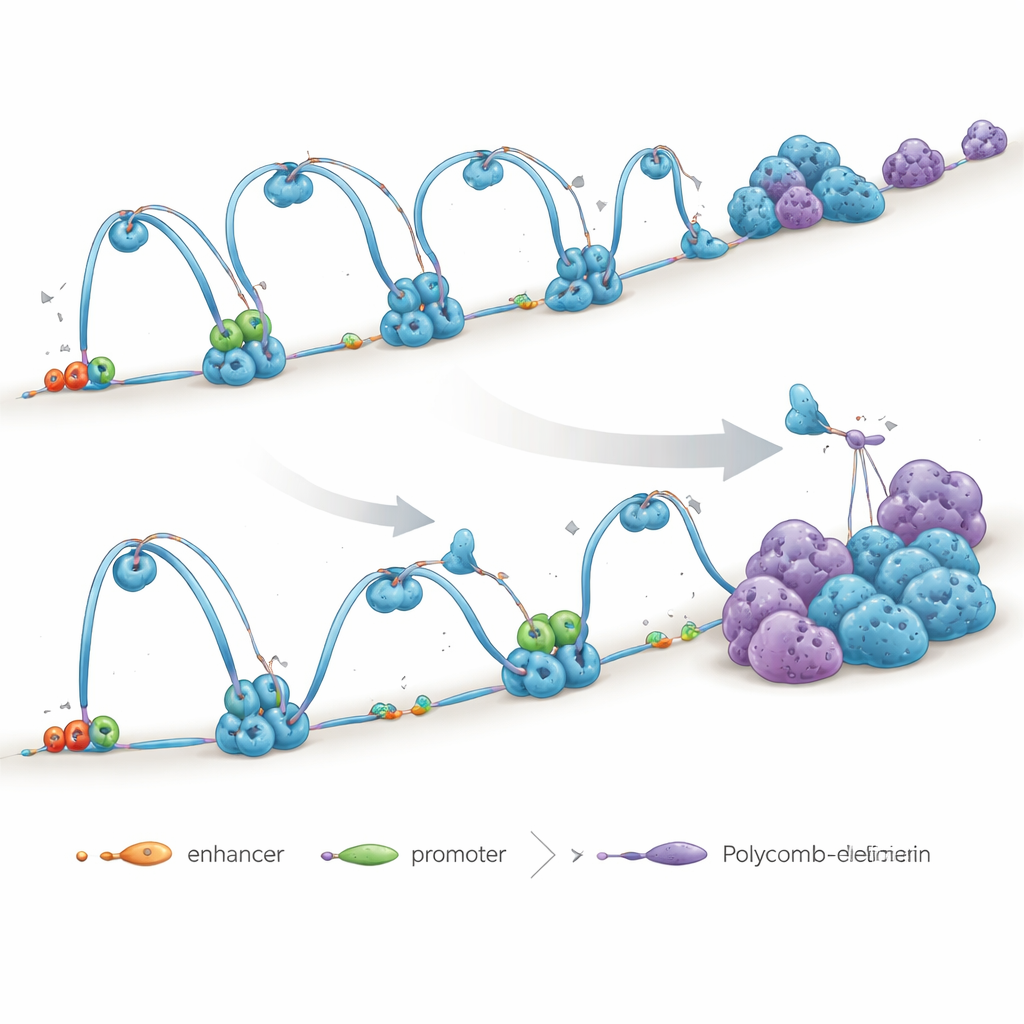
Autorzy zbadali także super-enhancery — duże skupiska elementów regulacyjnych działające jak „super-wyłączniki” genów definiujących tożsamość komórki. W normalnych komórkach te regiony są ze sobą powiązane i połączone z genami docelowymi przez pętle mediowane przez cohesin. Po utracie NIPBL znaczniki cohesin przy wielu z tych super-enhancerów zmalały, a pętle je łączące osłabły, podczas gdy niektóre długodystansowe kontakty między różnymi regionami super-enhancerów wzrosły w sposób nieuporządkowany. Sugeruje to, że gdy zwykłe obwody pętlowe się rozpadają, genom może rekompensować to przez formowanie szerszych, mniej precyzyjnych połączeń, co potencjalnie rozmywa ostry mechanizm włączania/wyłączania potrzebny genom rozwojowym.
Dlaczego to ma znaczenie dla rozwoju i chorób
W sumie badanie ukazuje NIPBL jako głównego organizatora genomu 3D podczas decyzji o losie komórkowym. Poprzez ładowanie i mobilizowanie cohesin, buduje i utrzymuje pętle, które pozwalają enhancerom i promotorom się komunikować, jednocześnie przeciwdziałając klastrowaniu regionów wyciszonych przez Polycomb. Gdy funkcja NIPBL jest upośledzona, pętle enhancer–promotor zapadają się, kompartmenty Polycomb się zacieśniają, a precyzyjnie zaplanowane programy genowe potrzebne do różnicowania trzustki zawodzą. Ten mechanistyczny obraz pomaga wyjaśnić, jak mutacje w genach związanych z cohesin mogą powodować złożone zespoły rozwojowe i daje wskazówki, jak subtelne zaburzenia składania genomu mogą przyczyniać się do chorób takich jak cukrzyca.
Cytowanie: Yu, L., Liu, Y., Zhang, J. et al. Impaired cohesin loading disrupts pancreatic differentiation by Polycomb-driven chromatin rewiring and loop collapse. Commun Biol 9, 590 (2026). https://doi.org/10.1038/s42003-026-09838-x
Słowa kluczowe: organizacja 3D genomu, cohesin i NIPBL, domeny Polycomb, różnicowanie trzustki, pętle enhancer–promotor