Clear Sky Science · ru
Анализ мигания флуоресценции в меченой ДНК с помощью скрытой модели Маркова
Почему важны крошечные вспышки света
В современных биологических лабораториях учёные часто наблюдают отдельные молекулы ДНК, присоединяя к ним флуоресцентную метку, которая моргает как микроскопический маяк. Эти вспышки несут богатую информацию о перемещении электронов по ДНК и о локальных изменениях окружения, но сигналы скрыты в шуме микроскопа и фона. В статье показано, как статистический инструмент машинного обучения — скрытая модель Маркова — может прорезать этот шум и выявить, когда метка действительно светится, когда она погашена и как долго длятся эти состояния, превращая беспорядочные световые сигналы в понятную физическую информацию.
Наблюдая за одной светящейся меткой на ДНК
Исследование сосредоточено на цепочках ДНК, меченных красным флуорофором (ATTO655) в определённом месте, вместе со специальной азотистой основой, способной захватывать заряд. Под постоянным лазерным облучением метка чередуется между испускающим «ВКЛ» состоянием и неиспускающим «ВЫКЛ». В состоянии ВКЛ краситель неоднократно поглощает фотоны и переизлучает их в виде флуоресценции. В состоянии ВЫКЛ электрон был передан, оставляя краситель в разделённой зарядовой конфигурации, которая не светится. Когда учёные регистрируют число фотонов, падающих на детектор в очень коротких временных интервалах — здесь по полмиллисекунды — получается неровная временная серия, в которой высокие и низкие счёты фотонов должны соответствовать периодам ВКЛ и ВЫКЛ, но сильно искажены случайными флуктуациями и фоновым светом.
Обучая модель «слушать» через шум
Для декодирования этих мерцающих траекторий авторы применяют скрытую модель Маркова (HMM) — структуру, хорошо известную в распознавании речи и финансах, но ещё недостаточно используемую в материаловедении. В этом случае скрытые состояния — просто ВКЛ и ВЫКЛ, а наблюдаемые данные — это числа фотонов в каждом временном бине. Команда предполагает, что при достаточном числе фотонов на бина для каждого состояния счёт можно аппроксимировать гладким колоколоподобным (гауссовским) распределением с разными средними. Используя байесовскую процедуру выборки, которая попеременно обновляет последовательность скрытых состояний и параметры, описывающие эти распределения и скорости переключения, HMM шаг за шагом определяет, какие сегменты траектории с наибольшей вероятностью соответствуют светящемуся или не светящемуся состоянию ДНК. В результате получается намного чище представленная двухуровневая последовательность состояний поверх шумной записи фотонного потока, а также оценочные вероятности переходов между ВКЛ и ВЫКЛ.
Измеряя длительности светлых и тёмных интервалов
Имея надёжную последовательность состояний, авторы собирают статистику по длительности эпизодов ВКЛ и ВЫКЛ. Они строят «графики мигания» — распределения вероятностей времени пребывания в каждом состоянии — и обнаруживают, что как длительности ВКЛ, так и ВЫКЛ следуют простому экспоненциальному убыванию. Из этих кривых они извлекают характерные времена релаксации: примерно 17,6 миллисекунды для состояния ВКЛ и 7,8 миллисекунды для состояния ВЫКЛ. По сравнению с внутренним процессом испускания отдельной молекулы красителя, происходящим на масштабе миллиардных долей секунды, эти интервалы в десятки миллисекунд чрезвычайно длинные. Состояние ВКЛ лучше понимать как квазастоционарный режим, в котором краситель совершает многие быстрые циклы поглощения–периспускания перед относительно редким переходом в ВЫКЛ. Длительный период ВЫКЛ указывает на удивительно стабильную зарядово-разделённую конфигурацию в системе ДНК–краситель, что подразумевает относительно медленную рекомбинацию заряда — возвращение в светящееся состояние.
Когда форма данных делает анализ успешным или нет
Интересно, что исследователи обнаружили: успех HMM сильно зависит от формы гистограммы подсчёта фотонов — распределения того, как часто встречается каждое число фотонов в бине. Когда гистограмма ясно показывает две вершины, одну для ВКЛ и одну для ВЫКЛ, модель восстанавливает чёткие последовательности состояний. Когда вершины сливаются в одну широкую гору, идентификация состояний становится гораздо более неоднозначной, хотя такие усреднённые величины, как среднее число фотонов и число событий, по-прежнему верно фиксируются. Команда показывает, что увеличение ширины временного бина обычно разделяет распределения ВКЛ и ВЫКЛ и даёт две пики, повышая устойчивость, но ценой потери информации о очень коротких событиях. Они предлагают практические эмпирические правила: минимальная надёжно измеримая длительность состояния составляет несколько крат выбранной ширины бина, а заметно бимодальная гистограмма — хороший индикатор того, что анализ заслуживает доверия.
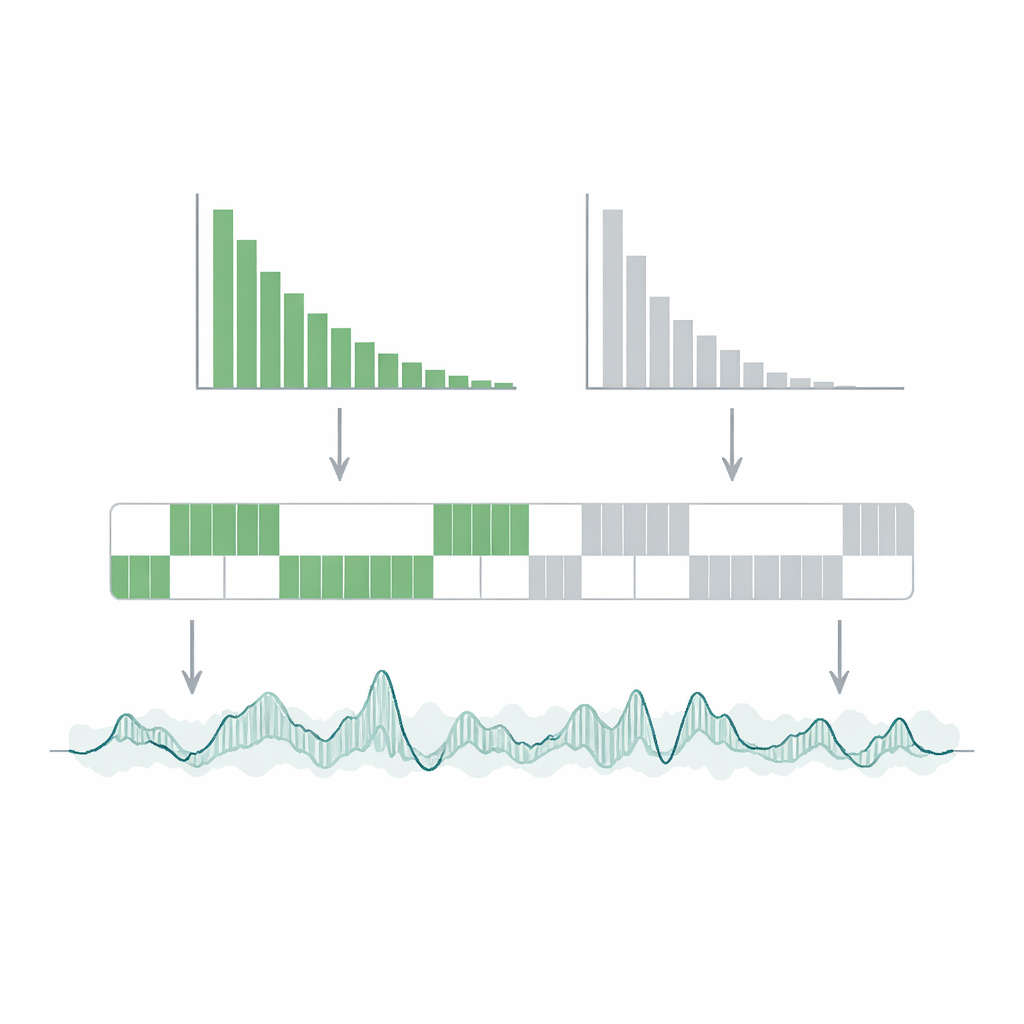
Что это значит для чтения молекулярных мерцаний
Объединив эксперименты по флуоресценции одиночных молекул с тщательно построенной скрытой марковской моделью, эта работа превращает шумное мигание из помехи в количественный зонд движения электронов вдоль ДНК. Вывод о том, что состояния ВЫКЛ длятся порядка восьми миллисекунд, показывает, что зарядово-разделённые состояния в этой конструкции ДНК–краситель необычно долговечны, тогда как примерно 18-миллисекундные периоды ВКЛ свидетельствуют о том, что перед каждой тёмной паузой может быть испущено большое число фотонов. Не менее важно то, что статья конкретно описывает, как такие выборы, как ширина временного бина и качество сигнала, определяют, надёжны ли такие анализы временных рядов, предлагая ясный контрольный список для будущих экспериментов. В совокупности эти достижения приближают исследователей к возможности считывать детальное электрическое и структурное поведение биомолекул непосредственно по их крошечным вспышкам света.
Цитирование: Furuta, T., Fan, S., Takada, T. et al. Hidden Markov model analysis of fluorescence blinking in fluorescently labeled DNA. Sci Rep 16, 11306 (2026). https://doi.org/10.1038/s41598-026-40876-x
Ключевые слова: флуоресценция одиночных молекул, электронный перенос в ДНК, мигание флуоресценции, скрытые марковские модели, подсчёт фотонов