Clear Sky Science · ru
Фосфорегуляция нового полускрывающегося (hemi-arrestin) MAPK-скэффолда Sms1 предотвращает преждевременное спаривание
Как дрожжи решают, пора ли спариваться
Одноклеточные грибы, такие как шизофилярные (fission) дрожжи, сталкиваются с фундаментальным выбором: продолжать деление или приостановить рост, чтобы поискать партнёра и слиться. Решиться на спаривание в неподходящий момент может быть смертельно опасно — особенно когда пищи предостаточно или подходящих партнёров мало. В этом исследовании обнаружен молекулярный «распределитель», белок Sms1, который помогает дрожжам определять, когда условия подходящие, включать программу спаривания на поверхности клетки и затем выключать её до того, как это станет вредным.
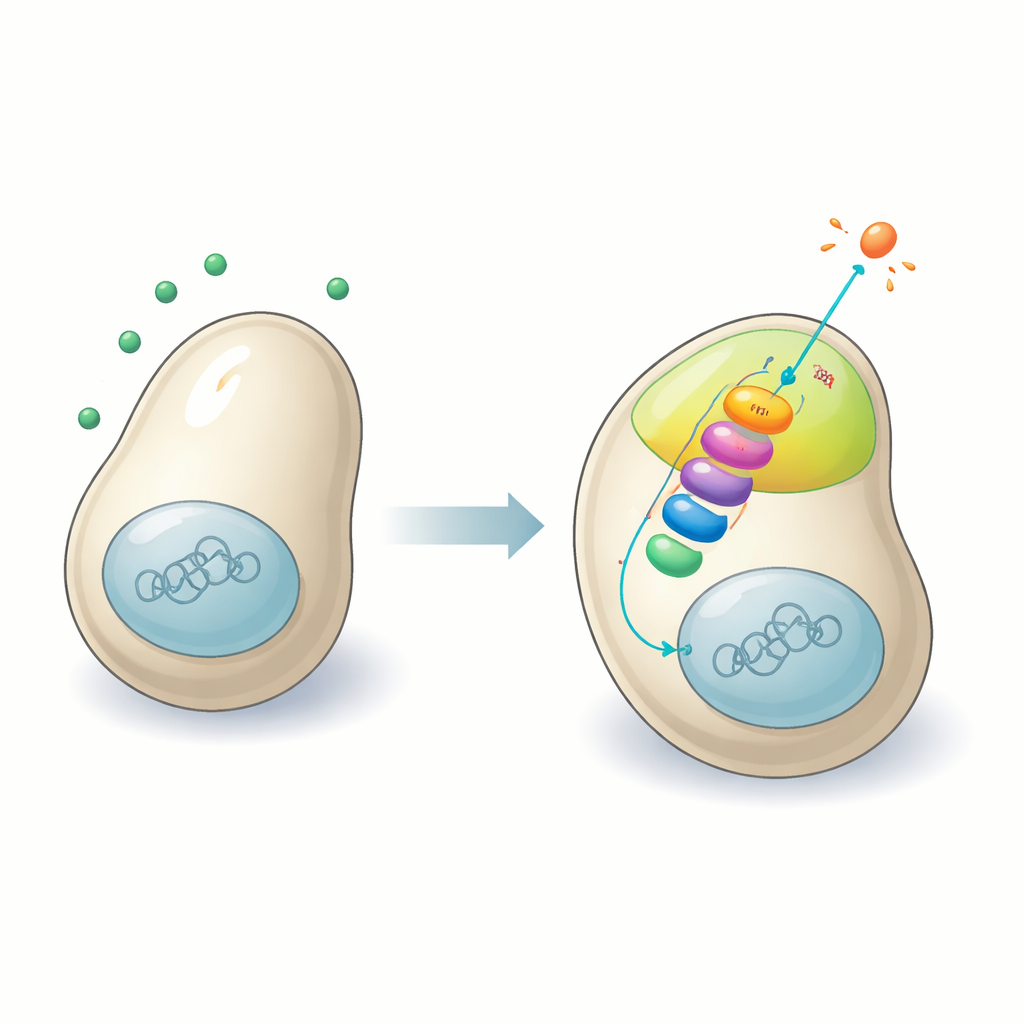
Молекулярный диалог на поверхности клетки
Клетки дрожжей общаются с потенциальными партнёрами с помощью феромонов — химических сигналов, обнаруживаемых рецепторами на поверхности клетки. Эти рецепторы связаны с хорошо известным внутриклеточным путём передачи сигналов — каскадом MAP-киназ, трёхступенчатой цепью белковых киназ, которые передают и усиливают сигналы. Во многих организмах специальные скэффолд-белки удерживают эти киназы в нужном месте, чтобы передача сигнала происходила эффективно. Долгое время учёные считали, что у шизофилярных дрожжей путь спаривания функционирует без такого скэффолда. Тщательно отслеживая белки и их взаимодействия, авторы опровергают это представление и выявляют Sms1 как недостающего организатора, который собирает компоненты пути спаривания вместе на плазматической мембране.
Создание сфокусированного центра спаривания
Когда клетка шизофилярных дрожжей улавливает феромон от совместимого партнёра, на её поверхности формируются небольшие, кратковременные «патчи полярности». Эти патчи — способ клетки прощупывать окружение и решать, в каком направлении расти к партнёру. Исследователи показывают, что Sms1 привлекается в эти патчи благодаря двум ключевым свойствам: арестиноподобному домену, который цепляется за специфические мембранные липиды, и прямому связыванию с субъединицей G-белка, активируемой феромонным рецептором. Оказавшись в патче, Sms1 действует как узел, физически ассоциируясь со всеми тремя звеньями каскада MAP-киназ. Это локальное стечение значительно усиливает сигнал, побуждающий клетку начать направленный рост и подготовку к слиянию.
От передачи сигнала к изменению формы
Изучая мутантные дрожжи без Sms1, исследователи обнаруживают, что спаривание фактически не происходит: критическая MAP-киназа Spk1 едва активируется, гены, необходимые для полового дифференцирования, не включаются должным образом, и клетки не формируют нормальные проекции для спаривания. Даже когда средняя киназа в цепи генетически запрограммирована быть постоянно активной, клеткам всё равно нужен Sms1, чтобы превратить общий сигнал в сфокусированный структурный ответ на поверхности клетки. Это указывает на то, что Sms1 делает не только «включение» сигнала: он помогает преобразовать этот сигнал в точные изменения формы клетки, гарантируя, что рост направлен на реального партнёра, а не случайно в пространство.
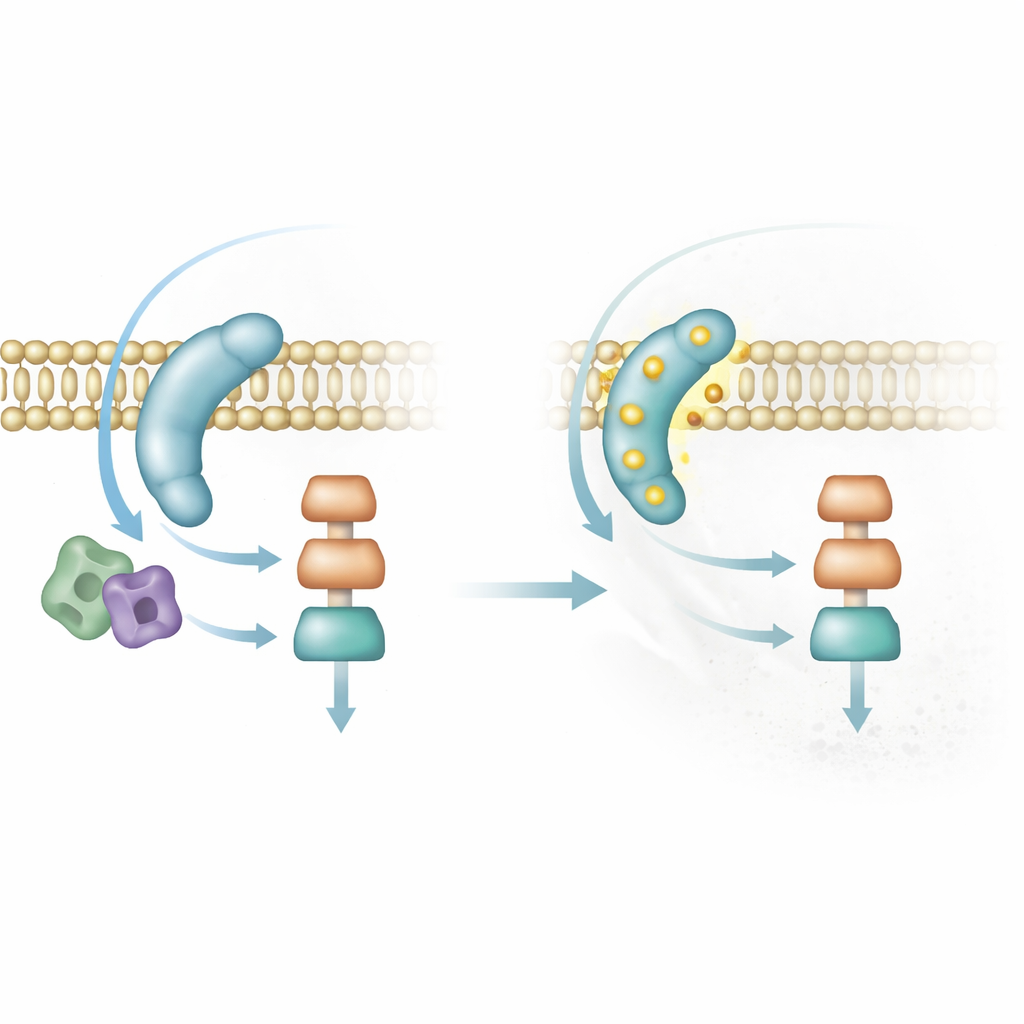
Встроенный тормоз, чтобы избежать неверного времени
Сигналы, включающие спаривание, должны также выключаться. Исследователи обнаружили, что Sms1 регулируется фосфорилированием — добавлением небольших химических меток киназами, включая ту самую MAP-киназу, которую он помогает активировать. Когда Sms1 фосфорилируется в определённых сайтах, он отталкивается от мембраны и сигнализационный патч растворяется. Дрожжи, сконструированные с нефосфорилируемой формой Sms1, накапливают долговечные яркие патчи, становятся чрезмерно чувствительны к феромонам и пытаются спариваться даже при избытке питательных веществ или после того, как уже слились в зиготу. Напротив, фосфо-имитирующая версия Sms1 не может достигать мембраны и делает клетки стерильными. Этот петлевой механизм обратной связи обеспечивает кратковременное и контекст-зависимое включение спаривания.
Что это означает за пределами дрожжей
Эта работа выявляет Sms1 как новый тип скэффолда MAP-киназы: он использует аррестиноподобную складку и гибкие участки, чтобы якорить сигнальные ферменты на поверхности клетки, а затем освобождать их, когда задача выполнена. Хотя структура Sms1 существенно отличается от известных скэффолдов у животных и печёночных (budding) дрожжей, он выполняет поразительно схожие функции — собирает каскад, локализует его в определённых мембранных участках и выключается фосфорилированием. Это указывает на то, что разнообразные организмы независимо пришли к сопоставимым решениям одной и той же проблемы: как строго контролировать мощные сигнальные пути, чтобы сложные решения, такие как спаривание, происходили только в нужном месте и в нужное время.
Цитирование: Sieber, B., Merlini, L., Li, W. et al. Phosphoregulation of the novel hemi-arrestin MAPK scaffold Sms1 prevents untimely mating. Nat Commun 17, 4084 (2026). https://doi.org/10.1038/s41467-026-70631-9
Ключевые слова: MAPK-сигнализация, спаривание дрожжей, скэффолд-белок, полярность клетки, обратная связь сигнала