Clear Sky Science · fr
La phosphorégulation du nouveau scaffold MAPK hémi-arrestine Sms1 empêche l’accouplement intempestif
Comment la levure décide qu’il est temps de s’accoupler
Les champignons unicellulaires comme la levure de fission font face à un choix fondamental : continuer à se diviser ou interrompre leur croissance pour chercher un partenaire et fusionner. S’engager dans l’accouplement au mauvais moment peut être mortel, en particulier lorsque la nourriture est abondante ou que les partenaires appropriés sont rares. Cette étude met au jour une protéine de « répartition » moléculaire, appelée Sms1, qui aide la levure à détecter quand les conditions sont favorables, à activer un programme d’accouplement à la surface cellulaire, puis à l’éteindre avant qu’il ne cause des dommages.
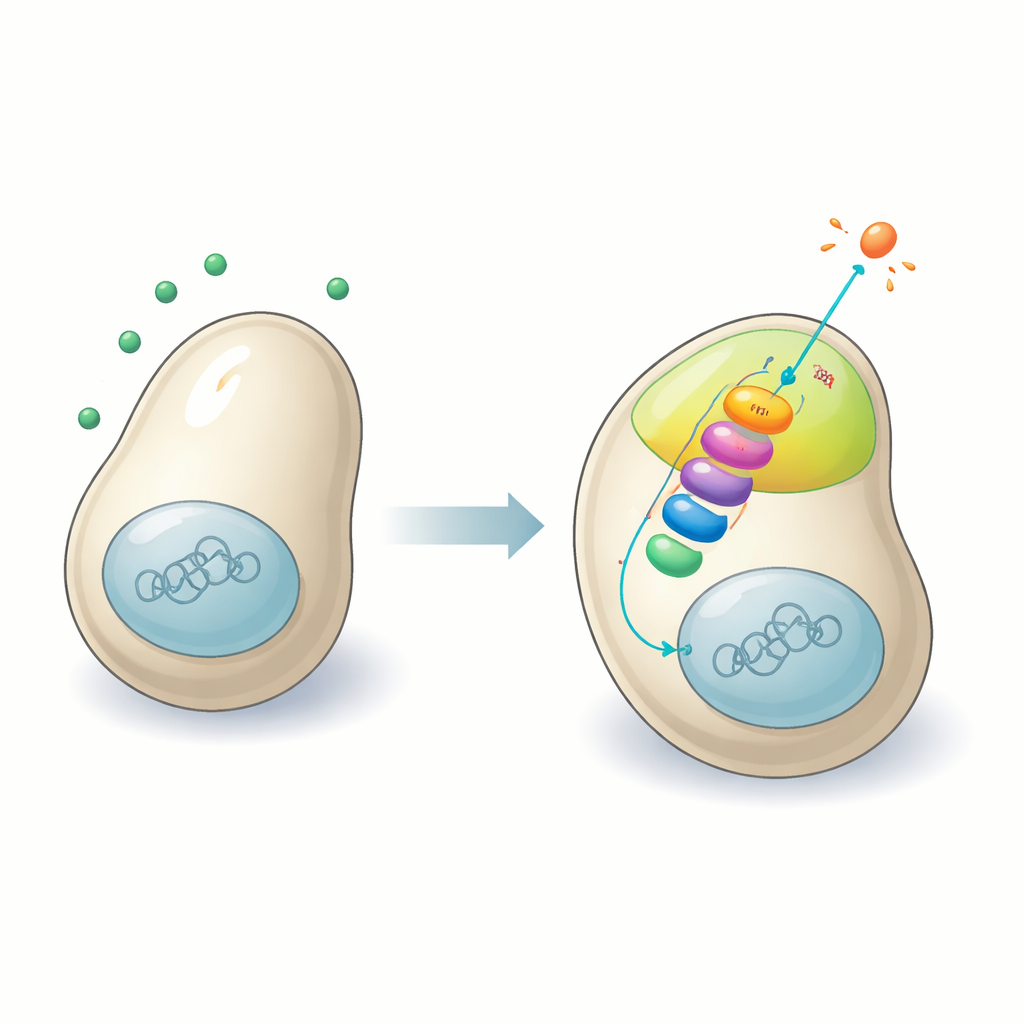
Une conversation moléculaire à la surface cellulaire
Les cellules de levure communiquent avec des partenaires potentiels à l’aide de phéromones — des signaux chimiques détectés par des récepteurs à la surface cellulaire. Ces récepteurs se relient à une voie de signalisation bien connue à l’intérieur des cellules appelée cascade de MAP kinases, une chaîne en trois étapes de kinases protéiques qui relaient et amplifient les signaux. Chez de nombreux organismes, des protéines échafaudage spéciales maintiennent ces kinases ensemble au bon endroit afin que les signaux soient transmis efficacement. Pendant des décennies, les scientifiques ont pensé que la levure de fission exécutait sa voie d’accouplement sans un tel échafaudage. En suivant soigneusement les protéines et leurs interactions, les auteurs renversent cette idée et révèlent Sms1 comme l’organisateur manquant qui rassemble les composants de la voie d’accouplement au niveau de la membrane plasmique.
Construire un point chaud d’accouplement focalisé
Lorsqu’une cellule de levure de fission détecte la phéromone d’un partenaire compatible, elle forme de petites « taches de polarité » transitoires à sa surface. Ces taches sont la manière dont la cellule sonde son environnement et décide où croître vers un partenaire. Les chercheurs montrent que Sms1 est recrutée vers ces taches grâce à deux caractéristiques clés : un domaine de type arrestine qui s’attache à des lipides membranaires spécifiques, et une liaison directe à une sous-unité de protéine G activée par le récepteur de la phéromone. Une fois au niveau de la tache, Sms1 agit comme un hub, s’associant physiquement aux trois niveaux de la cascade MAPK. Ce regroupement local intensifie fortement le signal qui indique à la cellule de lancer une croissance orientée et de se préparer à la fusion.
Du relais du signal au changement de forme
En examinant des levures mutantes dépourvues de Sms1, l’équipe constate que l’accouplement échoue essentiellement : la MAP kinase cruciale Spk1 est à peine activée, les gènes nécessaires à la différenciation sexuelle ne sont pas correctement induits, et les cellules ne forment pas de projections d’accouplement normales. Même lorsque la kinase intermédiaire de la chaîne est conçue pour être activée en permanence, les cellules ont encore besoin de Sms1 pour transformer le signal global en une réponse structurelle focalisée à la surface cellulaire. Cela indique que Sms1 fait plus que simplement activer le signal. Elle aide à traduire ce signal en changements précis de la forme cellulaire, en veillant à ce que la croissance soit dirigée vers un véritable partenaire plutôt qu’au hasard dans l’espace.
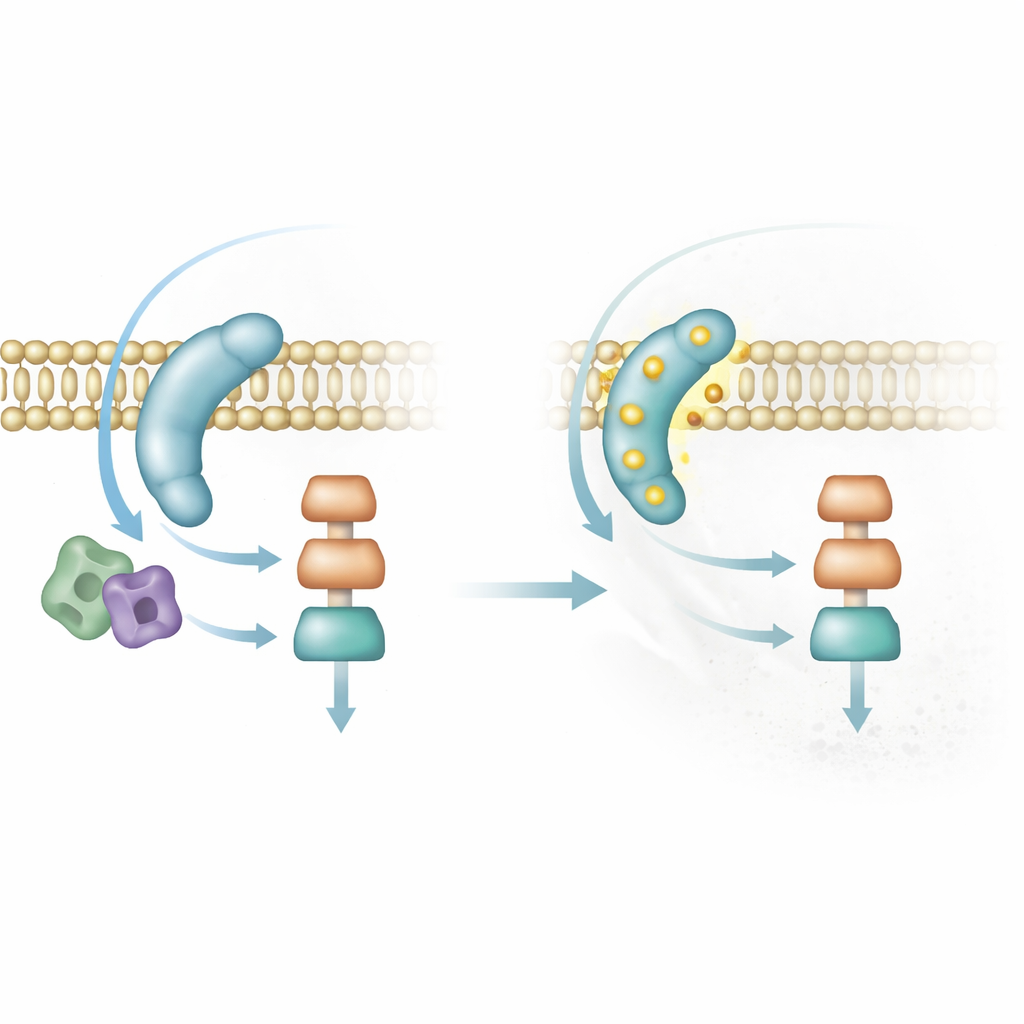
Un frein intégré pour éviter un mauvais timing
Les signaux qui déclenchent l’accouplement doivent aussi être éteints. Les chercheurs découvrent que Sms1 est contrôlée par phosphorylation — de petites marques chimiques ajoutées par des kinases, y compris la MAP kinase même qu’elle contribue à activer. Lorsque Sms1 est phosphorylée en des sites spécifiques, elle est repoussée de la membrane et la tache de signalisation se dissout. Des levures génétiquement modifiées pour porter une forme non phosphorylable de Sms1 accumulent des taches lumineuses et durables, deviennent excessivement sensibles aux phéromones et tentent de s’accoupler même lorsque les nutriments sont abondants ou après avoir déjà fusionné en zygote. À l’inverse, une version mimant la phosphorylation de Sms1 ne peut pas atteindre la membrane et rend les cellules stériles. Cette boucle de rétroaction garantit que l’accouplement n’est activé que brièvement et dans le bon contexte.
Ce que cela signifie au-delà de la levure
Ce travail révèle Sms1 comme un nouveau type d’échafaudage de MAP kinases : elle utilise un repli de type arrestine et des régions flexibles pour ancrer des enzymes de signalisation à la surface cellulaire, puis les libère une fois la tâche accomplie. Bien que Sms1 ait une structure très différente des échafaudages connus chez les animaux et la levure bourgeonnante, elle remplit des fonctions remarquablement similaires — assembler la cascade, la localiser à des sites membranaires spécifiques et être arrêtée par phosphorylation. Cela suggère que des organismes divers ont évolué indépendamment des solutions comparables au même problème : comment contrôler strictement des voies de signalisation puissantes afin que des décisions complexes comme l’accouplement n’aient lieu qu’au bon endroit et au bon moment.
Citation: Sieber, B., Merlini, L., Li, W. et al. Phosphoregulation of the novel hemi-arrestin MAPK scaffold Sms1 prevents untimely mating. Nat Commun 17, 4084 (2026). https://doi.org/10.1038/s41467-026-70631-9
Mots-clés: signalisation MAPK, accouplement de la levure, protéine échafaudage, polarité cellulaire, rétroaction du signal