Clear Sky Science · pt
Estereosseletividade e plasticidade funcional de um mesmo bolso de ligação a ligantes em TRPM3
Interruptor da dor em células nervosas
Por que algumas pessoas sentem mais dor que outras, e como uma molécula vegetal ou um medicamento contra epilepsia pode ajustar esse sinal para cima ou para baixo? Este estudo examina um minúsculo portão nas células nervosas, chamado TRPM3, que ajuda a detectar calor doloroso e é alterado em alguns transtornos do desenvolvimento neurológico. Ao observar exatamente como diferentes pequenas moléculas se encaixam no mesmo bolso desse portão, os pesquisadores revelam como detalhes químicos sutis podem ligar ou desligar o canal, com implicações claras para futuros tratamentos da dor e da epilepsia. 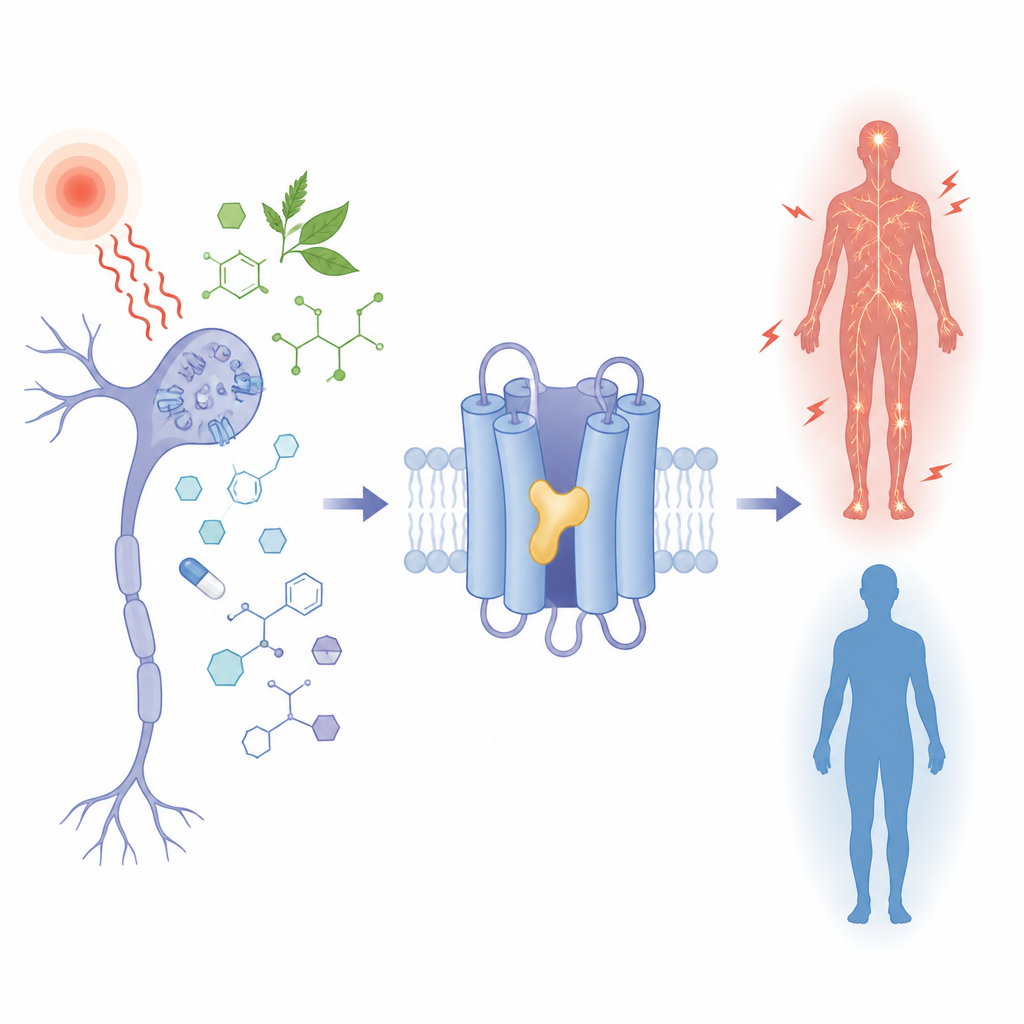
Um sensor de calor com um sítio de ancoragem compartilhado
TRPM3 é um canal iônico encontrado em nervos sensoriais que detectam calor nocivo e em várias células cerebrais. Quando está aberto, permite o fluxo de partículas carregadas, ajudando a transmitir sinais de dor. Em condições de doença ou quando o canal carrega certas alterações herdadas, esse fluxo torna-se excessivo e está ligado a crises convulsivas, atraso no desenvolvimento e sensibilidade alterada à dor. Vários compostos derivados de plantas e um antigo anticonvulsivante, primidona, são conhecidos por acalmar o TRPM3, enquanto uma molécula sintética chamada CIM0216 o ativa fortemente. Ainda assim, todas essas moléculas quimicamente diversas atuam na mesma parte do canal, e até agora não estava claro como um único bolso poderia abrigar tanto “freios” quanto “aceleradores”.
Imagens do bolso em alta resolução
A equipe usou criomicroscopia eletrônica, que pode visualizar proteínas individuais congeladas em uma fina camada de gelo, para capturar o TRPM3 sozinho e ligado a diferentes compostos. Eles se concentraram em uma cavidade formada por quatro hélices (S1–S4) e uma região próxima chamada domínio TRP, que juntas criam um bolso de ancoragem versátil. Mapas de alta resolução mostraram que primidona, dois compostos vegetais (isosakuranetina e ononetina) e CIM0216 ocupam posições sobrepostas nessa cavidade, mas situam-se em alturas e ângulos ligeiramente diferentes. A primidona se aloja no centro, os compostos vegetais se estendem mais em direção ao interior da célula, e o CIM0216 alcança mais a parte externa da célula. Essas poses distintas ditam quais aminoácidos no bolso contactam cada ligante e ajudam a explicar seus efeitos funcionais tão diversos. 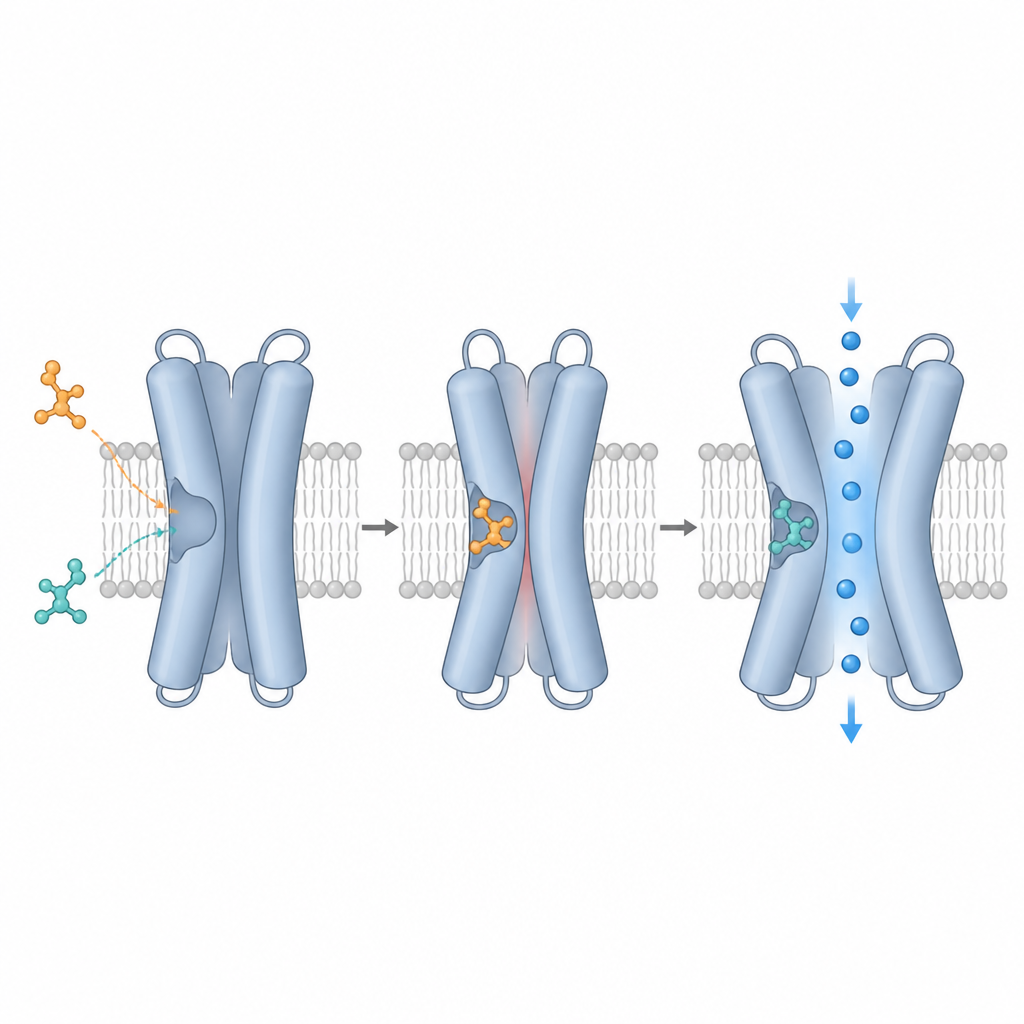
Fármacos imagem-espelhadas, efeitos opostos
Uma descoberta marcante é que o TRPM3 prefere fortemente uma das formas imagem-espelhadas, ou enantiômeros, de certos fármacos. A isosakuranetina comercial, derivada de plantas, mostrou-se uma mistura de dois enantiômeros. Ajustes estruturais e testes funcionais mostraram que apenas a forma R se encaixa firmemente no bolso e bloqueia o TRPM3 com potência, enquanto a forma S é em grande parte inativa no canal normal. O CIM0216 comporta-se de maneira semelhante: a forma R é um ativador poderoso e amplificador da atividade do canal, enquanto a forma S é bem mais fraca. Ao resolver a estrutura com R‑CIM0216 puro, os autores descobriram que esse ativador se projeta no espaço normalmente ocupado por uma cadeia lateral de tirosina específica, forçando-a a deslocar-se, um movimento que pode favorecer a abertura do canal.
Quando a resposta do portão se inverte
Para investigar como o bolso controla a função, os pesquisadores mudaram sistematicamente aminoácidos individuais que o revestem e depois mediram como as células respondiam aos vários ligantes. Algumas mutações enfraqueceram a ligação de forma generalizada; outras afetaram seletivamente apenas um composto, dependendo de onde esse ligante se situava na cavidade. Notavelmente, algumas alterações não apenas modificaram a potência, mas inverteram o efeito: em um mutante, o R‑CIM0216 normalmente ativador passou a reduzir a atividade basal, enquanto a S‑isosakuranetina, geralmente inativa, tornou-se um ativador potente. Esses resultados mostram que o bolso é funcionalmente plástico: pequenas variações na forma do canal ou na estereoquímica do ligante podem alternar o canal entre os estados “ligado” e “desligado”, mesmo que o mesmo sítio físico seja usado.
Mutações de pacientes e desafios no tratamento
O estudo também descreve dois pacientes com epilepsia que carregam variantes raras de TRPM3 que se localizam diretamente nesse bolso. Essas alterações tornam o canal mais ativo em repouso e mais facilmente acionável, consistente com um efeito de ganho de função. Crucialmente, as variantes também tornam o canal muito menos sensível à primidona e aos antagonistas vegetais, mesmo quando subunidades normais e mutantes estão misturadas, como ocorreria em um paciente. Isso sugere que, para tais indivíduos, doses padrão de primidona podem não ser suficientes para conter o TRPM3, ressaltando a necessidade de fármacos sob medida que considerem o bolso alterado.
O que isso significa para medicamentos futuros
Em conjunto, o trabalho revela o bolso de ligação de ligantes do TRPM3 como um centro de controle altamente adaptável, onde fármacos imagem-espelhadas e pequenas alterações na forma do canal podem alternar entre potencializar e bloquear sinais relacionados à dor. Para os projetistas de fármacos, essa flexibilidade é ao mesmo tempo um desafio e uma oportunidade: novas terapias para dor e transtornos cerebrais ligados ao TRPM3 precisarão considerar não apenas se um composto cabe no bolso, mas exatamente como sua forma 3D e mutações locais do canal direcionam o portão para configurações abertas ou fechadas.
Citação: Bazeli, B., Shkumatov, A.V., Schenck, S. et al. Stereoselectivity and functional plasticity of a common ligand-binding pocket in TRPM3. Nat Commun 17, 4556 (2026). https://doi.org/10.1038/s41467-026-71226-0
Palavras-chave: TRPM3, canal iônico, estereosseletividade, sinalização da dor, transtornos do desenvolvimento neurológico