Clear Sky Science · en
Stereoselectivity and functional plasticity of a common ligand-binding pocket in TRPM3
Pain switch in nerve cells
Why do some people feel more pain than others, and how might a plant molecule or epilepsy drug dial that signal up or down? This study looks at a tiny gate in nerve cells, called TRPM3, that helps sense painful heat and is altered in some neurodevelopmental disorders. By seeing exactly how different small molecules plug into the same pocket on this gate, the researchers reveal how subtle chemical details can turn the channel on or off, with clear implications for future pain and epilepsy treatments. 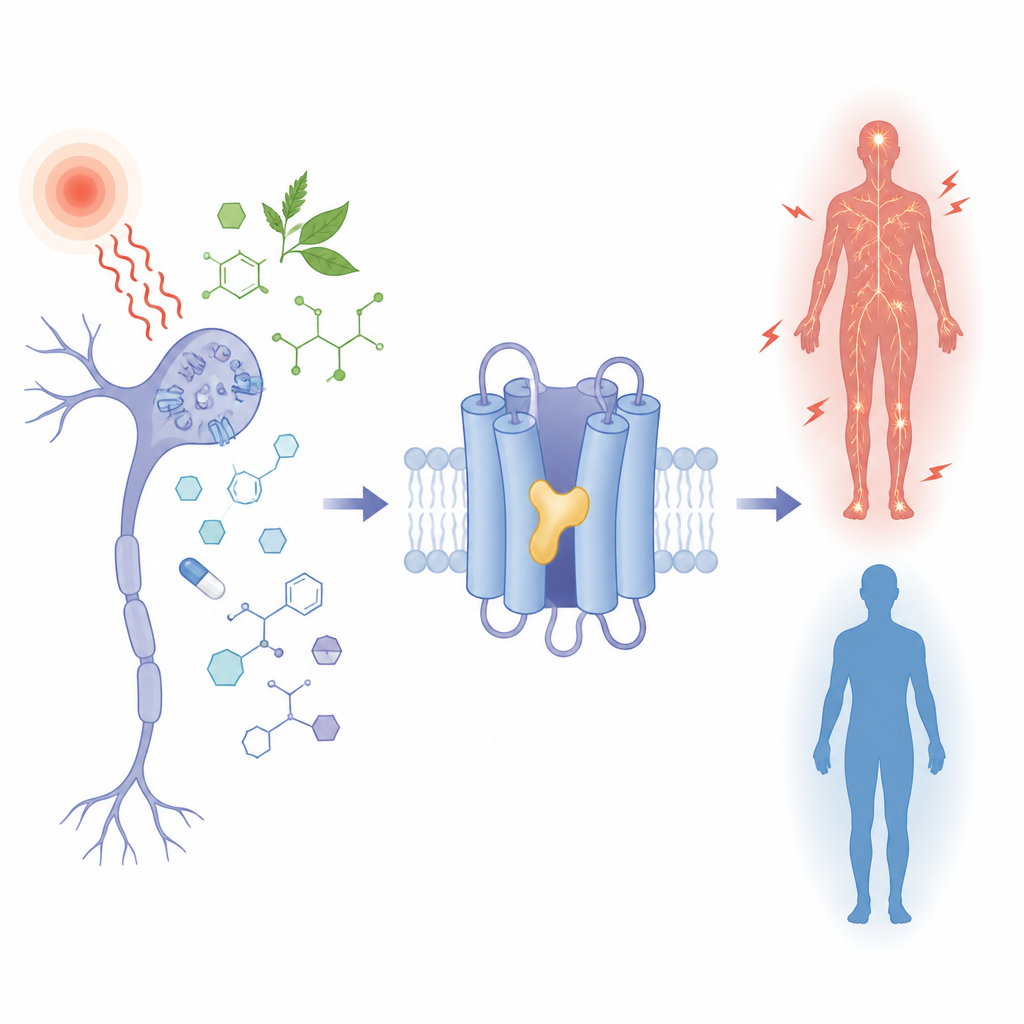
A heat sensor with a shared docking site
TRPM3 is an ion channel found in sensory nerves that detect noxious heat and in various brain cells. When open, it lets charged particles flow, helping to send pain signals. Under disease conditions or when the channel carries certain inherited changes, this flow becomes excessive and is linked to seizures, developmental delay, and altered pain sensitivity. Several plant-derived compounds and a long-used epilepsy drug, primidone, are known to calm TRPM3, while a synthetic molecule called CIM0216 strongly activates it. Yet all of these chemically diverse molecules act on the same part of the channel, and until now it was unclear how a single pocket could host both “brakes” and “accelerators.”
Imaging the pocket in high detail
The team used cryo-electron microscopy, which can visualize individual proteins frozen in a thin layer of ice, to capture TRPM3 alone and bound to different compounds. They focused on a cavity formed by four helices (S1–S4) and a nearby region called the TRP domain, which together create a versatile docking pocket. High-resolution maps showed that primidone, two plant compounds (isosakuranetin and ononetin), and CIM0216 all occupy overlapping positions in this cavity but sit at slightly different heights and angles. Primidone nestles in the center, the plant compounds extend further toward the cell interior, and CIM0216 reaches more toward the outside of the cell. These distinct poses dictate which amino acids in the pocket contact each ligand and help explain their very different functional effects. 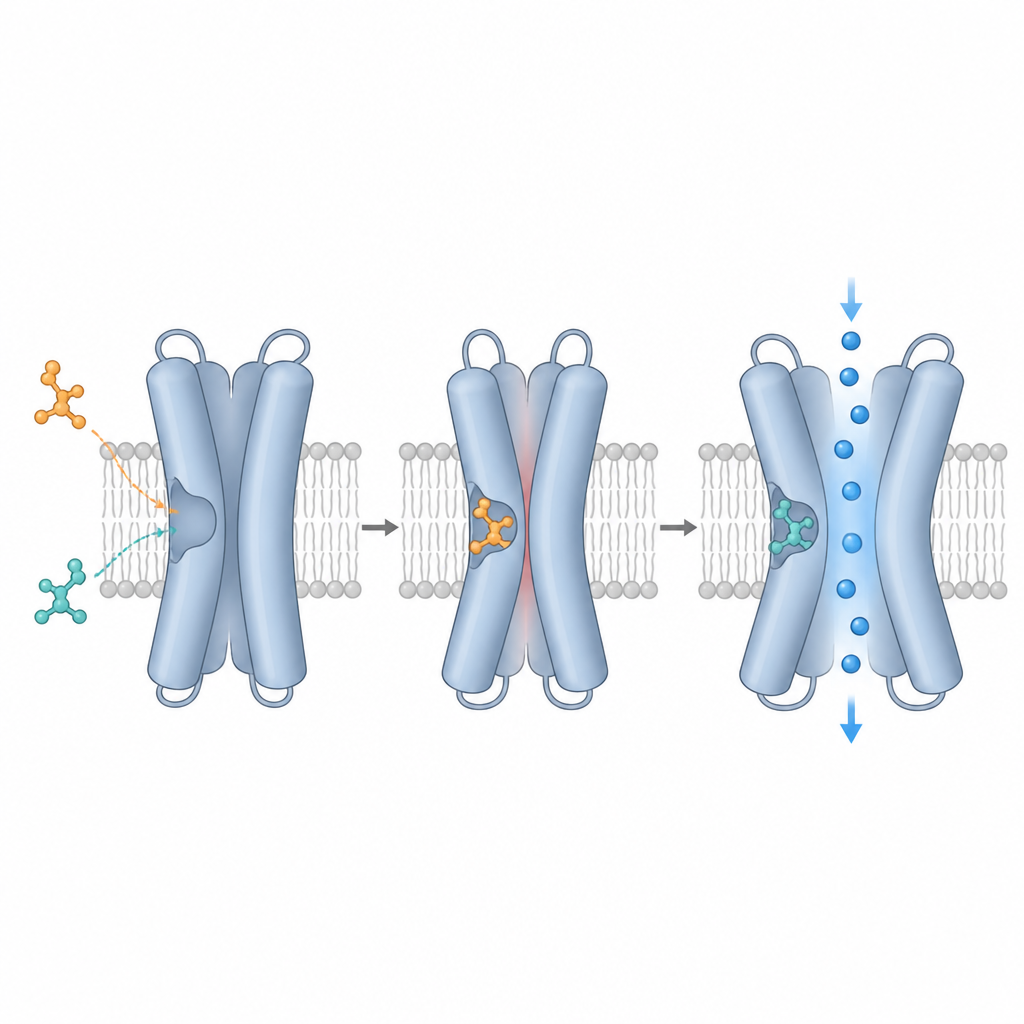
Mirror-image drugs, opposite effects
A striking finding is that TRPM3 strongly prefers one mirror-image form, or enantiomer, of certain drugs. Commercial isosakuranetin, derived from plants, turned out to be a mix of two enantiomers. Structural fits and functional tests showed that only the R-form tightly matches the pocket and potently blocks TRPM3, while the S-form is largely inactive on the normal channel. CIM0216 behaves similarly: the R-form is a powerful activator and booster of channel activity, whereas the S-form is far weaker. By solving the structure with pure R‑CIM0216, the authors discovered that this activator pushes into space normally occupied by a specific tyrosine side chain, forcing it to swing aside, a motion that may help promote channel opening.
When the gate’s response flips
To probe how the pocket controls function, the researchers systematically changed individual amino acids that line it and then measured how cells responded to the various ligands. Some mutations weakened binding across the board; others selectively affected only one compound, depending on where that ligand sat in the cavity. Remarkably, a few changes did not merely alter potency but reversed the effect: in one mutant, the normally activating R‑CIM0216 instead dampened basal activity, while the usually inactive S‑isosakuranetin became a strong activator. These results show that the pocket is functionally plastic: small shifts in channel shape or ligand stereochemistry can flip the channel between “on” and “off” states, even though the same physical site is used.
Patient mutations and treatment challenges
The study also describes two patients with epilepsy carrying rare TRPM3 variants that sit directly in this pocket. These changes make the channel more active at rest and more easily triggered, consistent with a gain-of-function effect. Crucially, the variants also make the channel far less sensitive to primidone and to the plant antagonists, even when normal and mutant subunits are mixed as they would be in a patient. This suggests that for such individuals, standard primidone doses may not sufficiently rein in TRPM3, underscoring the need for tailored drugs that take the altered pocket into account.
What this means for future medicines
Together, the work reveals TRPM3’s ligand-binding pocket as a highly adaptable control hub where mirror-image drugs and tiny tweaks in channel shape can switch between boosting and blocking pain-related signals. For drug designers, that flexibility is both a challenge and an opportunity: new therapies for pain and TRPM3-linked brain disorders will need to consider not just whether a compound fits the pocket, but exactly how its 3D shape and local channel mutations steer the gate toward open or closed configurations.
Citation: Bazeli, B., Shkumatov, A.V., Schenck, S. et al. Stereoselectivity and functional plasticity of a common ligand-binding pocket in TRPM3. Nat Commun 17, 4556 (2026). https://doi.org/10.1038/s41467-026-71226-0
Keywords: TRPM3, ion channel, stereoselectivity, pain signaling, neurodevelopmental disorders