Clear Sky Science · fr
Sélectivité stéréochimique et plasticité fonctionnelle d’un même site de liaison aux ligands dans TRPM3
Interrupteur de la douleur dans les cellules nerveuses
Pourquoi certaines personnes ressentent-elles plus la douleur que d’autres, et comment une molécule d’origine végétale ou un médicament anti‑épileptique peut‑il régler ce signal à la hausse ou à la baisse ? Cette étude examine une minuscule porte dans les cellules nerveuses, appelée TRPM3, qui contribue à détecter la chaleur douloureuse et est altérée dans certains troubles du neurodéveloppement. En observant précisément comment différentes petites molécules se logent dans la même poche de cette porte, les chercheurs montrent comment des détails chimiques subtils peuvent activer ou désactiver le canal, avec des implications claires pour de futurs traitements de la douleur et de l’épilepsie. 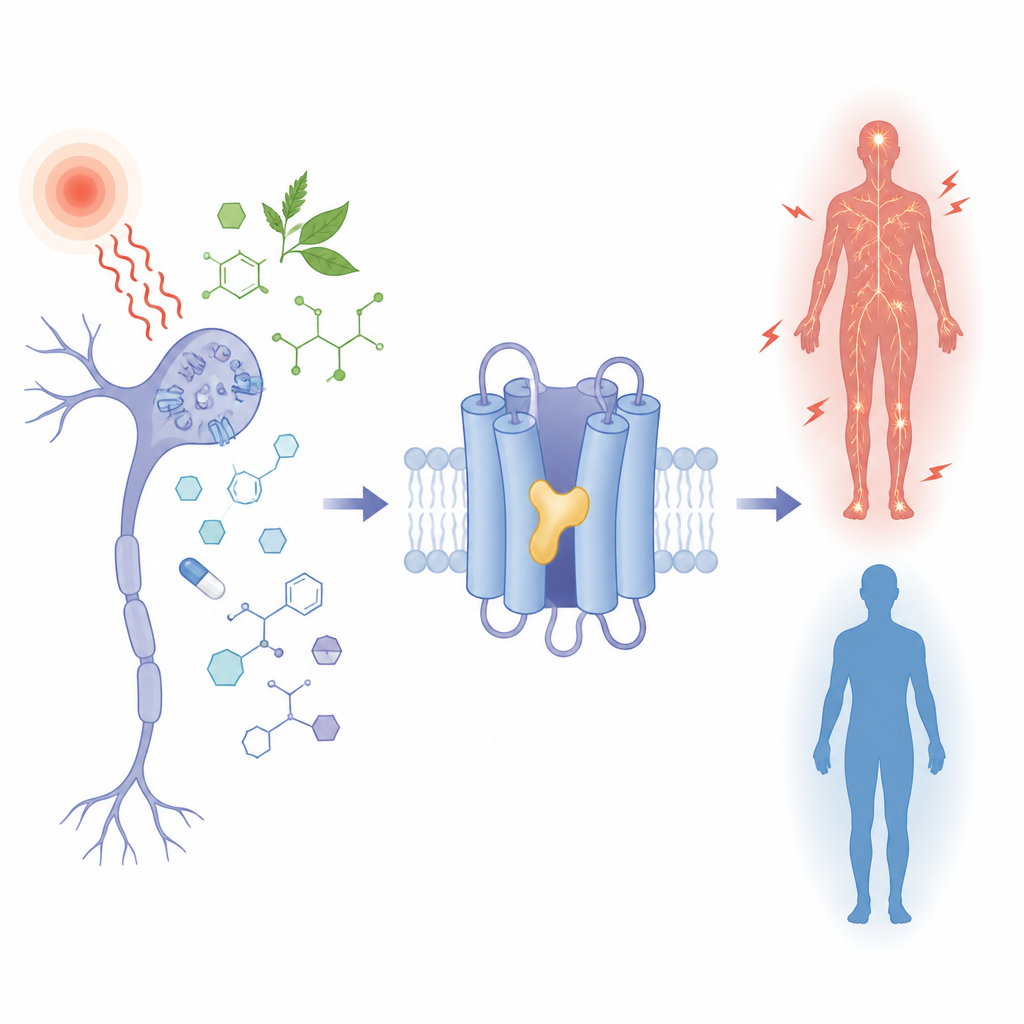
Un capteur de chaleur avec un site d’ancrage partagé
TRPM3 est un canal ionique présent dans les nerfs sensoriels qui détectent la chaleur nocive et dans diverses cellules cérébrales. Lorsqu’il est ouvert, il laisse passer des particules chargées, contribuant à la transmission des signaux de douleur. En cas de maladie ou lorsque le canal porte certaines variations héréditaires, ce flux devient excessif et est lié à des crises, des retards de développement et une sensibilité à la douleur modifiée. Plusieurs composés d’origine végétale et un médicament anti‑épileptique utilisé de longue date, la primidone, calment TRPM3, tandis qu’une molécule synthétique appelée CIM0216 l’active fortement. Pourtant, toutes ces molécules chimiquement diverses agissent sur la même région du canal, et jusqu’ici il n’était pas clair comment une seule poche pouvait accueillir à la fois des « freins » et des « accélérateurs ».
Imager la poche en haute résolution
L’équipe a utilisé la cryo‑microscopie électronique, qui permet de visualiser des protéines individuelles congelées dans une fine couche de glace, pour capturer TRPM3 seul et lié à différents composés. Ils se sont concentrés sur une cavité formée par quatre hélices (S1–S4) et une région proche appelée domaine TRP, qui ensemble créent une poche d’ancrage polyvalente. Les cartes à haute résolution ont montré que la primidone, deux composés végétaux (l’isosakuranétine et l’ononétine) et le CIM0216 occupent tous des positions qui se chevauchent dans cette cavité, mais se placent à des hauteurs et des angles légèrement différents. La primidone se niche au centre, les composés végétaux s’étendent davantage vers l’intérieur de la cellule, et le CIM0216 atteint plus vers l’extérieur. Ces poses distinctes dictent quelles acides aminés de la poche entrent en contact avec chaque ligand et aident à expliquer leurs effets fonctionnels très différents. 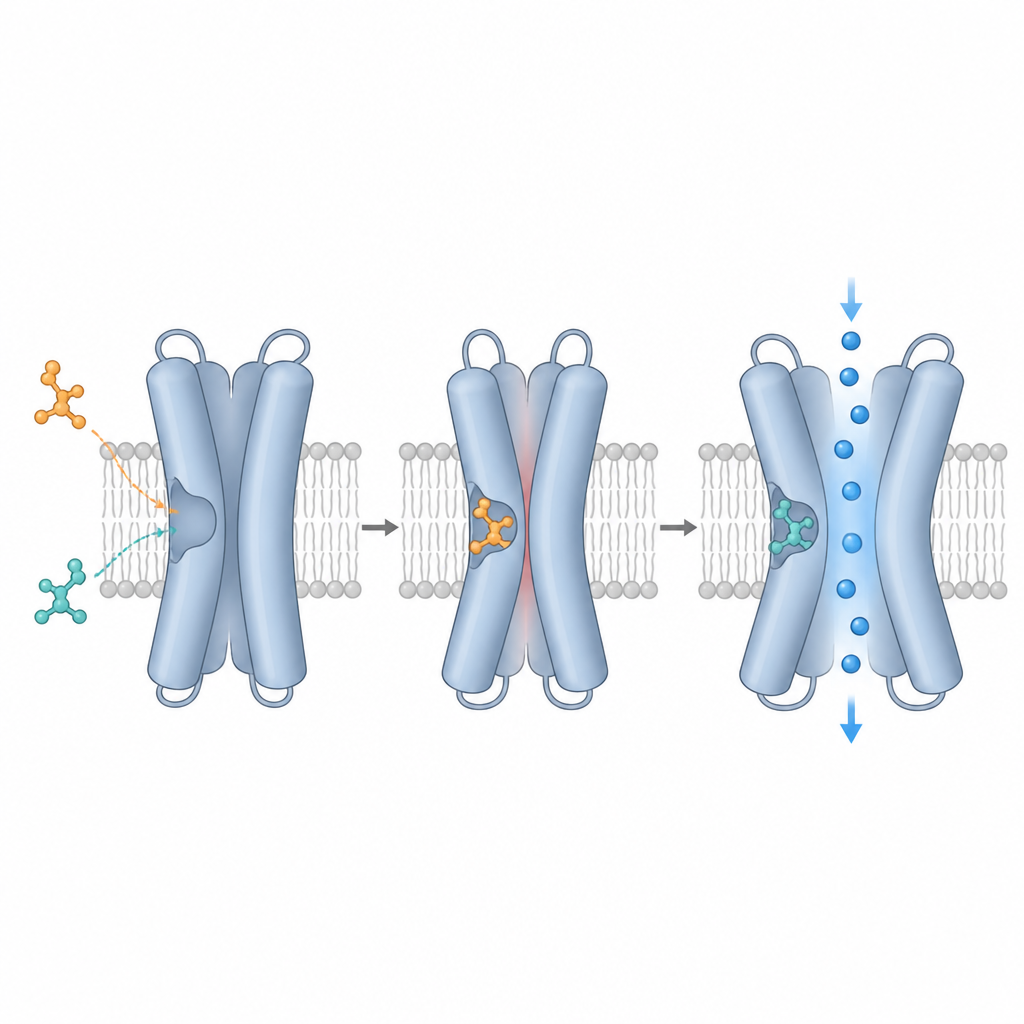
Médicaments images miroir, effets opposés
Une découverte marquante est que TRPM3 préfère nettement l’une des formes images miroir, ou énantiomères, de certains médicaments. L’isosakuranétine commerciale, d’origine végétale, s’est révélée être un mélange des deux énantiomères. Les ajustements structuraux et les tests fonctionnels ont montré que seul l’énantiomère R s’accorde étroitement avec la poche et bloque puissamment TRPM3, tandis que la forme S est largement inactive sur le canal normal. Le CIM0216 présente un comportement similaire : la forme R est un activateur puissant et augmente fortement l’activité du canal, alors que la forme S est beaucoup plus faible. En résolvant la structure avec le R‑CIM0216 pur, les auteurs ont découvert que cet activateur s’insère dans l’espace normalement occupé par la chaîne latérale d’une tyrosine spécifique, la forçant à se décaler, un mouvement qui pourrait favoriser l’ouverture du canal.
Quand la réponse de la porte s’inverse
Pour explorer comment la poche contrôle la fonction, les chercheurs ont modifié systématiquement des acides aminés individuels qui la bordent, puis mesuré la réponse cellulaire aux différents ligands. Certaines mutations ont affaibli la liaison de façon globale ; d’autres ont affecté sélectivement un seul composé, selon l’endroit où ce ligand se trouvait dans la cavité. De manière remarquable, quelques changements n’ont pas seulement modifié la puissance, mais ont inversé l’effet : dans un mutant, le R‑CIM0216 normalement activateur a en fait atténué l’activité basale, tandis que l’isosakuranétine S habituellement inactive est devenue un fort activateur. Ces résultats montrent que la poche est fonctionnellement plastique : de petits déplacements dans la conformation du canal ou dans la stéréochimie du ligand peuvent basculer le canal entre états « ouvert » et « fermé », bien que le même site physique soit utilisé.
Mutations patientes et défis thérapeutiques
L’étude décrit également deux patients épileptiques portant de rares variantes de TRPM3 situées directement dans cette poche. Ces changements rendent le canal plus actif au repos et plus facilement déclenché, cohérent avec un effet de gain de fonction. De manière cruciale, les variantes rendent aussi le canal beaucoup moins sensible à la primidone et aux antagonistes d’origine végétale, même lorsque des sous‑unités normales et mutantes sont mélangées comme cela se produirait chez un patient. Cela suggère que, pour ces individus, les doses habituelles de primidone peuvent ne pas suffire à réprimer TRPM3, soulignant la nécessité de médicaments adaptés qui tiennent compte de la poche altérée.
Ce que cela signifie pour les médicaments futurs
Dans l’ensemble, ce travail révèle la poche de liaison aux ligands de TRPM3 comme un centre de contrôle hautement adaptable où des médicaments images miroir et de petites modifications de la forme du canal peuvent commuter entre l’amplification et le blocage des signaux liés à la douleur. Pour les concepteurs de médicaments, cette flexibilité est à la fois un défi et une opportunité : de nouvelles thérapies pour la douleur et les troubles cérébraux associés à TRPM3 devront prendre en compte non seulement si un composé s’insère dans la poche, mais aussi comment sa forme 3D et les mutations locales du canal orientent la porte vers des configurations ouvertes ou fermées.
Citation: Bazeli, B., Shkumatov, A.V., Schenck, S. et al. Stereoselectivity and functional plasticity of a common ligand-binding pocket in TRPM3. Nat Commun 17, 4556 (2026). https://doi.org/10.1038/s41467-026-71226-0
Mots-clés: TRPM3, canal ionique, sélectivité stéréochimique, signalisation de la douleur, troubles du neurodéveloppement