Clear Sky Science · pl
Strukturalne podstawy eksportu galaktanu związane z lipidami przez mykobakteryjny transporter ABC Wzm-Wzt
Dlaczego ta bakteryjna brama ma znaczenie
Leki, które leczą gruźlicę i pokrewne zakażenia, muszą przedrzeć się przez jedne z najtrwalszych ścian komórkowych w przyrodzie. Mykobakterie, do których należy czynnik wywołujący gruźlicę, otaczają się grubą, woskową zbroją. W centrum tej zbroi znajduje się długi oparty na cukrach szkielet zwany arabinogalaktanem. Niniejsze badanie ujawnia, w niemal atomowym detalu, jak kluczowa maszyna molekularna — bramka eksportowa Wzm-Wzt — przesuwa ważny element budulcowy tego szkieletu przez błonę komórkową. Zrozumienie tego procesu otwiera nowe możliwości osłabienia ściany bakteryjnej i projektowania przyszłych antybiotyków.
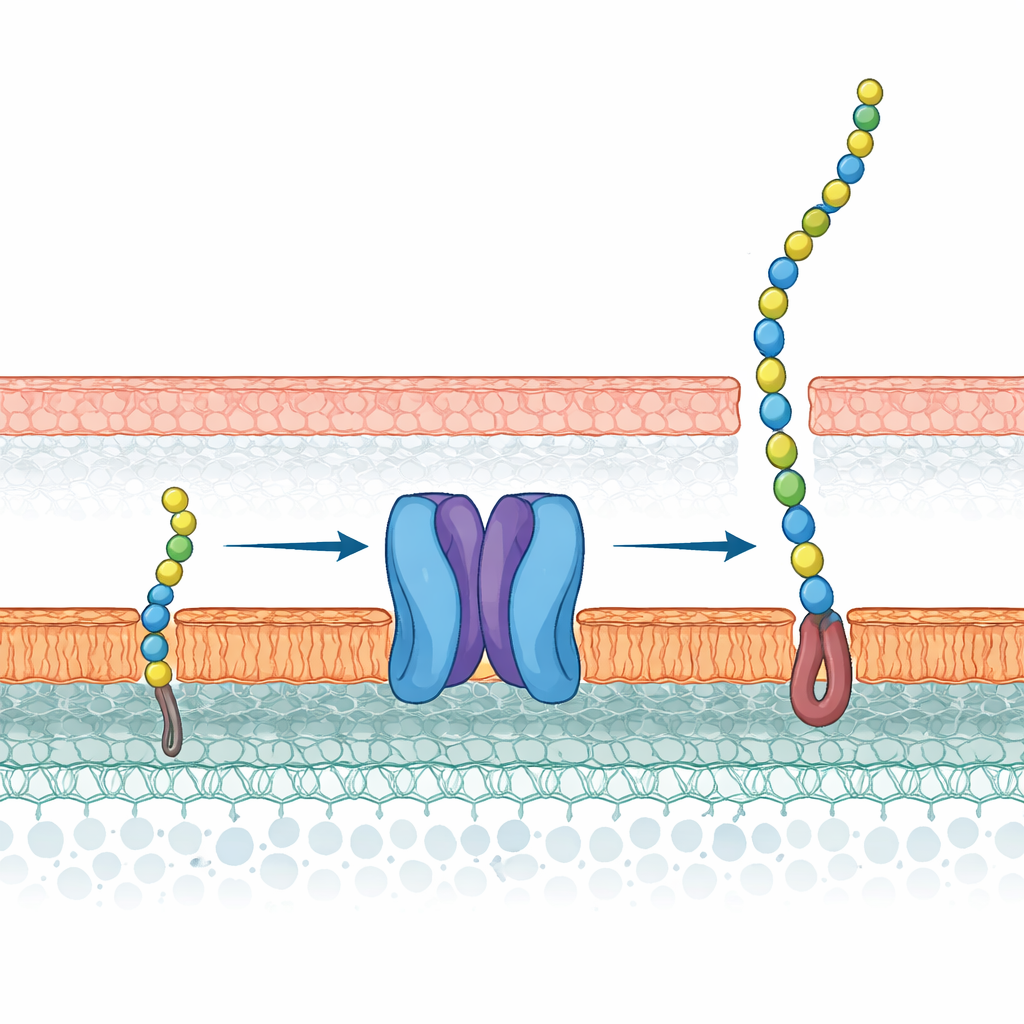
Szczególna zbroja mykobakterii
Większość bakterii ma otoczki komórkowe zbudowane z warstw lipidów i cząsteczek cukrowych, ale mykobakterie idą w tej kwestii dalej. Ich błona wewnętrzna jest pokryta gęstą siecią cukrową — arabinogalaktanem — który następnie jest ozdobiony bardzo długimi kwasami tłuszczowymi, tworząc ochronną zewnętrzną „mykomembranę”. Kilka pierwszoliniowych leków przeciwgruźliczych już celuje w enzymy budujące części tej macierzy. Zanim jednak te enzymy będą mogły działać, bakteria musi odwrócić prekursor zwany galaktanem związanym z lipidami z wnętrza komórki na zewnętrzną powierzchnię błony wewnętrznej. Ten prekursor łączy tłusty ogon zanurzony w błonie z długim łańcuchem cukrowym galaktanu — co czyni go jednocześnie dużym i chemicznie niewygodnym do przemieszczania.
Odnalezienie maszyny eksportującej cukry
Wcześniejsze prace identyfikowały Wzm-Wzt jako transporter wykonujący to trudne przewrócenie. Podobnie jak inne transportery ABC, Wzm-Wzt zużywa paliwo komórkowe (ATP) w swoich częściowych domenach cytosolowych, aby wywoływać zmiany kształtu w kanale osadzonym w błonie. Wciąż jednak nie było jasne, jak taka maszyna mogłaby chwycić cząsteczkę będącą częścią tłuszczu, częściowo silnie naładowaną łącznikiem i częściowo masywnym łańcuchem cukrowym, a następnie przesuwać ją stopniowo przez błonę nie naruszając bariery komórkowej. Aby to rozwiązać, autorzy oczyścili Wzm-Wzt z patogenu Mycobacterium abscessus, osadzili go w detergencie lub w małych syntetycznych dyskach błonowych i użyli krioelektronowej mikroskopii, by uchwycić wiele migawkowych stanów transportera podczas jego cyklu pracy.
Migawki pracy molekularnej bramy
Struktury ukazują Wzm-Wzt jako parowany kanał w błonie połączony z dwoma napędzanymi przez ATP silnikami wewnątrz komórki. Wewnątrz kanału trzy ułożone jak pasy warstwy aromatycznych aminokwasów wyściełają potencjalną ścieżkę dla łańcucha cukrowego. Mały fragment białka po stronie cytosolowej, nazwany helisą bramkową, porusza się dramatycznie między pozycjami otwartą i zamkniętą w miarę wiązania i rozkładu ATP. Dodając syntetyczny odpowiednik naturalnego galaktanu związanego z lipidem, badacze zaobserwowali gęstość zgodną z cząsteczką zaklinowaną między dwoma helisami, z jej hydrofobowym ogonem wchodzącym najpierw do jamy i głową cukrową ustawioną przy wylocie kanału. Wspiera to tryb załadunku „najpierw lipid”, gdzie tłusty ogon służy jako uchwyt rozpoznawany przez transporter, zanim łańcuch cukrowy zostanie przepchnięty dalej.
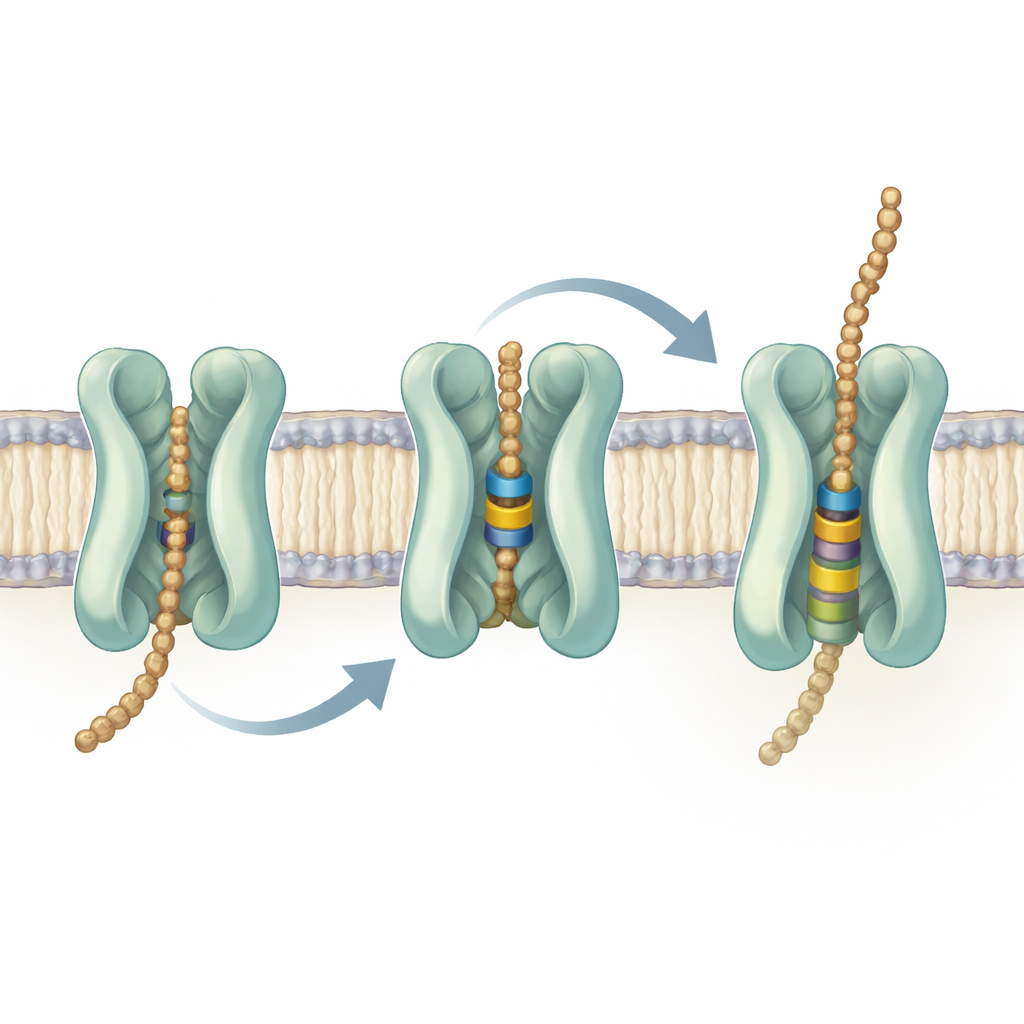
Testowanie ruchomych elementów bramy
Aby sprawdzić, które części Wzm-Wzt są niezbędne, zespół wprowadził precyzyjne mutacje i zbadał ich skutki w modelowym gatunku mykobakterii. Użyli genetycznego wyłącznika, by częściowo wyłączyć rodzimy transporter, a następnie dostarczyli normalne lub zmienione wersje z plazmidu. Gdy Wzm-Wzt działał, bakterie dobrze rosły i budowały prawidłowe ściany komórkowe. Gdy kluczowe reszty w miejscu wiązania ATP lub cała helisa bramkowa zostały zaburzone, komórki przestały rosnąć, akumulowały prekursory lipidowe i nadmiernie produkowały inne składniki ściany, które normalnie przyłączają się do arabinogalaktanu — to sygnatury przerwanego etapu eksportu. Mutacje w pobliskiej pętli przy wejściu do kanału również unieruchomiły transport, podczas gdy zmiany niektórych aromatycznych reszt głęboko w zatoce powodowały tylko częściowe spowolnienia. Te funkcjonalne testy, skojarzone ze strukturami, podkreślają helisę bramkową i tzw. pętlę LG jako aktywne elementy pomagające chwytać i przeskokowo przesuwać łańcuch cukrowy.
Nowe słabe miejsce w ścianie gruźlicy
Podsumowując, wyniki wspierają model, w którym galaktan związany z lipidami dokuje ogonem najpierw, przesuwa się między dwiema helisami, a następnie jego długi łańcuch cukrowy jest nawlekany przez wąski tunel wyłożony aromatycznymi resztami, podczas gdy ruchy napędzane ATP helisy bramkowej i pętli wejściowej wyciągają go na zewnątrz. Ponieważ składanie arabinogalaktanu jest niezbędne dla przeżycia mykobakterii, a Wzm-Wzt jest bardzo podatny na uszkodzenia, transporter ten wyróżnia się teraz jako obiecujący cel lekowy. Małe cząsteczki blokujące zatokę wiążącą lipidy lub zamrażające ruchome elementy bramki mogłyby zatrzymać konstrukcję ściany komórkowej i, w połączeniu z istniejącymi terapiami, pomóc przezwyciężyć oporne zakażenia mykobakteryjne.
Cytowanie: Garaeva, A.A., Fabianová, V., Savková, K. et al. Structural basis of lipid-linked galactan export by the mycobacterial ABC transporter Wzm-Wzt. Nat Commun 17, 2745 (2026). https://doi.org/10.1038/s41467-026-70429-9
Słowa kluczowe: gruźlica, ściana komórkowa bakterii, transporter ABC, arabinogalaktan, cele antybiotykowe