Dlaczego nasza skóra ciemnieje po ekspozycji na światło słoneczne lub pod wpływem pewnych hormonów i jak pęcherzyki pigmentu wewnątrz komórek wiedzą, kiedy „włączyć” produkcję barwnika? To badanie ujawnia zaskakującą rozmowę między dwiema drobnymi strukturami w komórkach pigmentowych — mitochondriami, „elektrowniami” komórki, i melanosomami, przedziałami, które wytwarzają i przechowują melaninę. Obserwując te struktury, gdy stykają się i rozchodzą w czasie rzeczywistym, badacze pokazują, jak krótkotrwałe kontakty pomagają ustawić właściwe wewnętrzne warunki do bezpiecznej i efektywnej syntezy melaniny.
Małe fabryki pigmentu wewnątrz naszych komórek
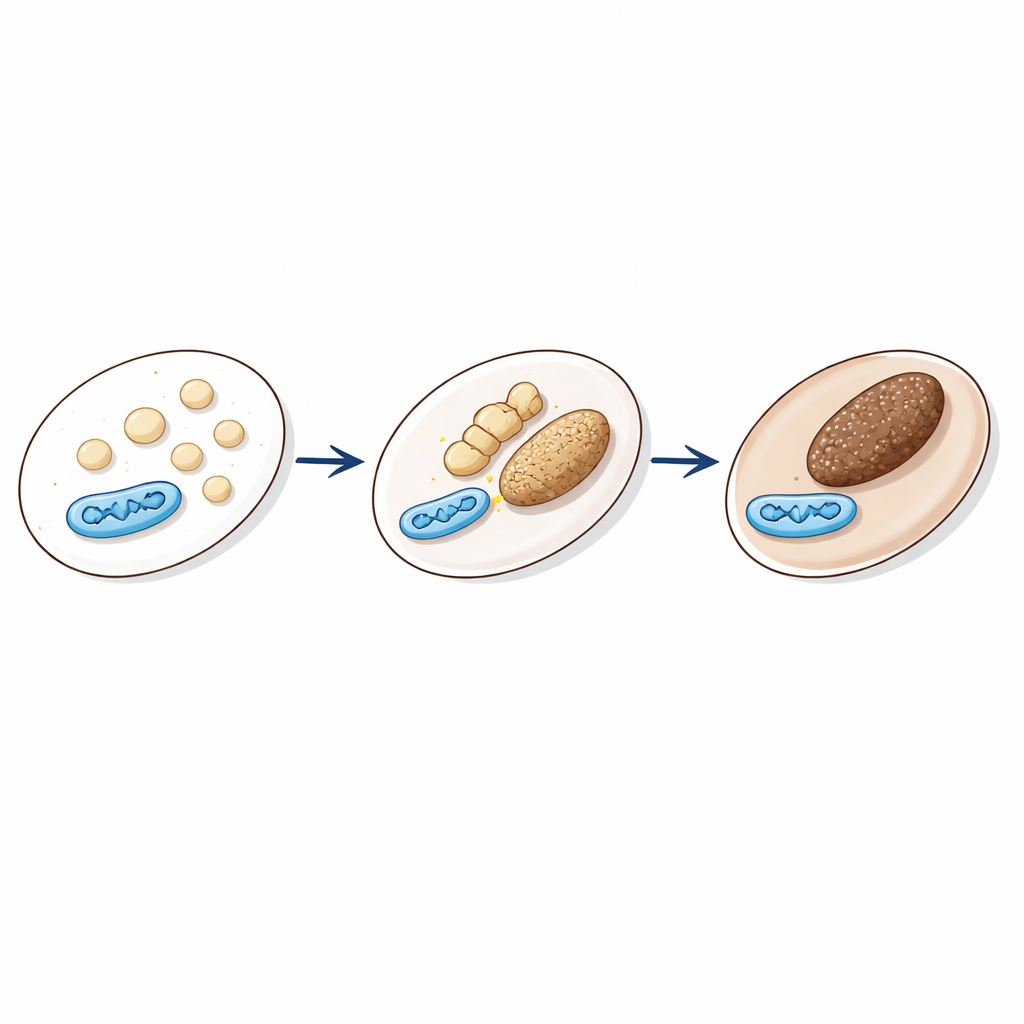
Melanina, barwnik nadający kolor skóry, włosom i oczom, jest wytwarzana i magazynowana w wyspecjalizowanych przedziałach zwanych melanosomami. Struktury te rozwijają się etapami, od jasnych, pustych łupin do ciemnych ziaren wypełnionych melaniną, które mogą być transportowane ku powierzchni komórki. Ich aktywność jest regulowana sygnałami, takimi jak hormon α‑MSH, którego poziom rośnie po ekspozycji na promieniowanie ultrafioletowe i który zwiększa pigmentację. Aby melanosomy działały poprawnie, chemia wewnątrz nich — w szczególności kwasowość i poziom wapnia — musi zmieniać się we właściwym czasie. Jednym z wczesnych kroków jest utworzenie białkowego rusztowania z włókien PMEL, co wymaga kwaśnego wnętrza. Później przedział staje się mniej kwaśny, aby enzymy produkujące melaninę mogły działać. To, jak te precyzyjne zmiany są zasilane i synchronizowane, było dotąd niejasne.
Gdy elektrownie spotykają ziarna pigmentu Figure 1.
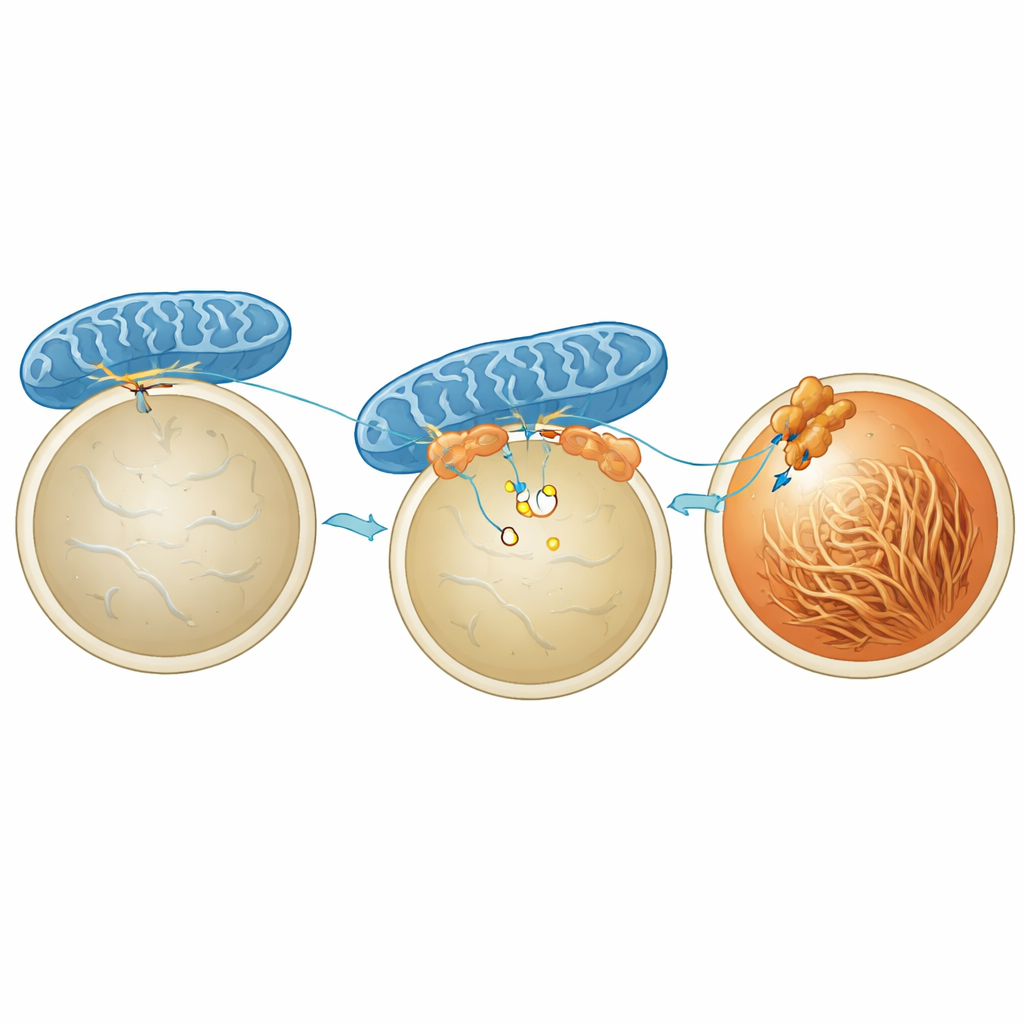
Zespół skupił się na miejscach fizycznego kontaktu między mitochondriami a melanosomami. Takie „uściski dłoni” organelli są ważne także w innych częściach komórki, na przykład między mitochondriami a retikulum endoplazmatycznym. Tu badacze opracowali system raportujący w żywych komórkach o nazwie MiMSBiT, który świeci, gdy mitochondria i melanosomy zbliżają się na tyle, że dwie zaprojektowane fragmenty białek mogą się połączyć. Stosując to narzędzie w komórkach czerniaka myszy, odkryli, że α‑MSH i pokrewne sygnały wywołują silny, lecz przejściowy wzrost kontaktów mitochondria–melanosom. Kontakty te osiągały szczyt około trzech godzin po stymulacji — w tym samym oknie czasowym, gdy melanosomy stawały się najbardziej kwaśne i gdy formowały się włókna PMEL — co sugeruje, że fizyczna bliskość obu organelli jest ściśle powiązana z dojrzewaniem ziaren pigmentu.
Zespół spinający: STIM1 i Mitofusin 2
Aby zrozumieć, co faktycznie trzyma mitochondria i melanosomy razem, naukowcy skoncentrowali się na białku MFN2, znanym już z łączenia mitochondriów z innymi organellami. Zmniejszenie ekspresji MFN2 w komórkach pigmentowych znacząco ograniczyło kontakt wywoływany hormonem i osłabiło wzrost pigmentacji, bez zmiany poziomów ani podstawowej aktywności enzymów produkujących melaninę. Kluczowym graczem po stronie melanosomu okazał się STIM1, szerzej znany jako czujnik wapnia w innym przedziale komórkowym. Używając metody znakowania bliskości i wysokorozdzielczego obrazowania, badacze pokazali, że pula STIM1 siedzi na melanosomach i krótkotrwale wiąże się z MFN2 na mitochondriach pod wpływem α‑MSH. Interakcja ta jest wyzwalana przez krótkotrwały spadek wapnia wewnątrz światła melanosomu, co powoduje skupianie się STIM1 i jego uchwyt MFN2, tworząc fizyczny most.
Dostawa energii i zakwaszanie wewnątrz melanosomów Figure 2.
Co zyskuje się przez przyciągnięcie mitochondriów do melanosomów? Badanie pokazuje, że te kontakty lokalnie zwiększają dostępność ATP, waluty energetycznej komórki, bezpośrednio przy powierzchni melanosomu. Używając fluorescencyjnego sensora ATP umieszczonego w błonie melanosomu, autorzy stwierdzili, że α‑MSH podnosi stężenie ATP wokół melanosomów w sposób zależny od produkcji energii przez mitochondria, ale niezależny od rozkładu cukrów w cytoplazmie. Gdy MFN2 lub STIM1 były zredukowane, ten lokalny wzrost ATP znikał, mimo że ogólna liczba kontaktów czy podstawowy metabolizm komórki nie ulegały dramatycznym zmianom. Dodatkowe ATP wydaje się zasilać pompy protonowe w błonie melanosomu, które aktywnie wtłaczają protony, tymczasowo zakwaszając wnętrze. Ten impuls kwasowości z kolei sprzyja składaniu się PMEL w uporządkowane włókna, które tworzą rusztowanie, na którym później można bezpiecznie osadzić melaninę.
Od kontaktów komórkowych do pigmentacji całego organizmu
Aby sprawdzić, czy ten mikroskopijny mechanizm ma znaczenie u żywych organizmów, badacze potraktowali embriony zebrafish lekiem zakłócającym ten sam region MFN2 potrzebny do wiązania STIM1. Rozwijające się ryby miały znacznie jaśniejsze ciała, co potwierdza, że zakłócenie tych kontaktów organelli upośledza normalną pigmentację in vivo. Razem wyniki układają się w etapową opowieść: sygnały hormonalne wywołują zmiany wapnia w niedojrzałych melanosomach; to aktywuje STIM1, który łączy się z MFN2, spinając mitochondria; mitochondria dostarczają wtedy ATP tam, gdzie jest potrzebne, aby zakwasić melanosomy i uporządkować rusztowanie PMEL; i dopiero wtedy może postępować intensywna produkcja melaniny. Dla laika oznacza to, że kolor, który widzimy na poziomie skóry i włosów, zależy od precyzyjnie zsynchronizowanych, nanoskalowych interakcji między maleńkimi strukturami głęboko wewnątrz każdej komórki pigmentowej.
Cytowanie: Shiiba, I., Ishikawa, Y., Oshio, H. et al. STIM1-Mitofusin2 interactions tether mitochondria and melanosome contacts that promote melanosome maturation.
Nat Commun17, 3593 (2026). https://doi.org/10.1038/s41467-026-70282-w
Słowa kluczowe: kontakt mitochondria–melanosom, produkcja melaniny, komunikacja organelli, pigmentacja skóry, komórkowa energia i ATP