Warum dunkelt unsere Haut nach Sonnenlicht oder bestimmten Hormonen nach, und wie wissen Pigmentgranula in Zellen, wann sie aktiv werden sollen? Diese Studie enthüllt ein unerwartetes Gespräch zwischen zwei winzigen Strukturen in Pigmentzellen — Mitochondrien, die „Kraftwerke“ der Zelle, und Melanosomen, die Kompartimente, in denen Melanin gebildet und gespeichert wird. Indem die Forschenden beobachteten, wie diese Strukturen sich in Echtzeit berühren und wieder trennen, zeigen sie, wie diese kurzen Kontakte die richtigen inneren Bedingungen schaffen, damit Melanin sicher und effizient aufgebaut werden kann.
Kleine Pigmentfabriken in unseren Zellen
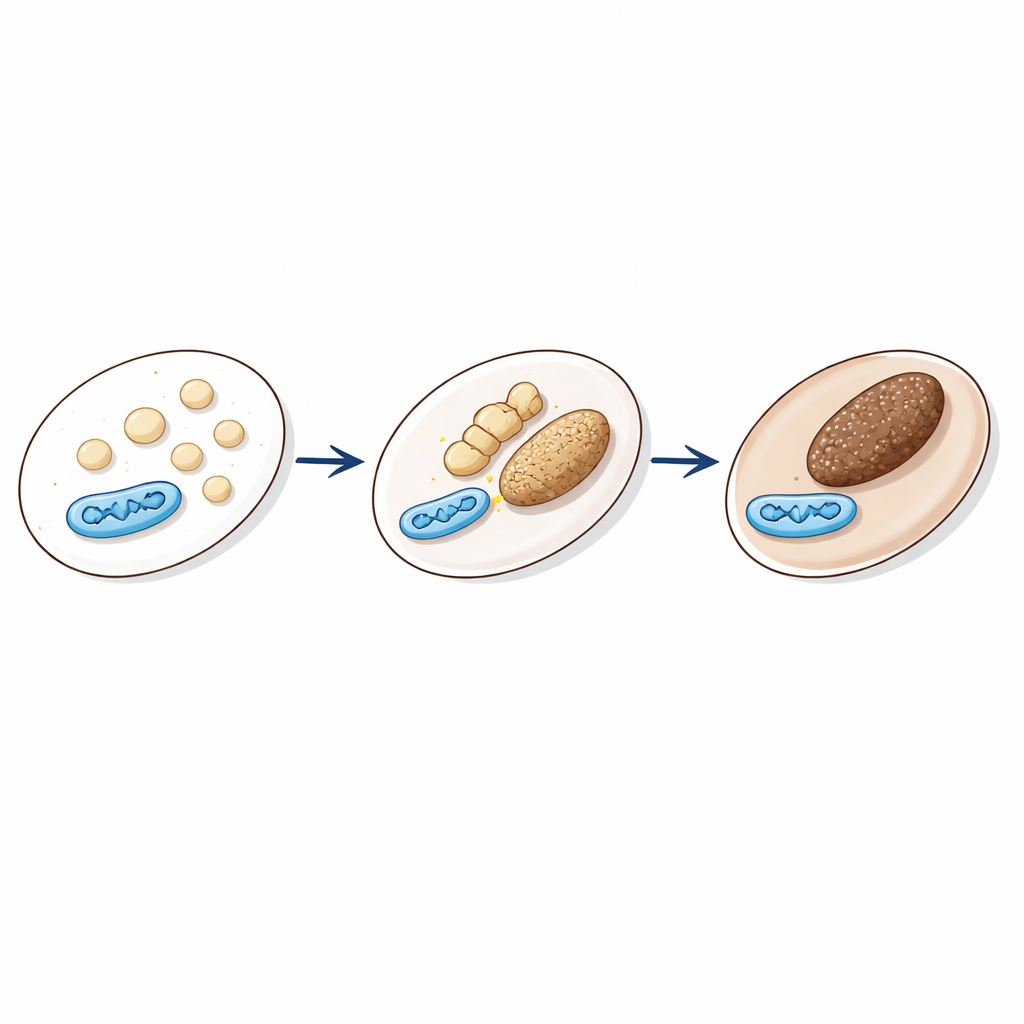
Melanin, das Pigment, das unsere Haut, Haare und Augen färbt, wird in spezialisierten Kompartimenten hergestellt und gespeichert, den Melanosomen. Diese Strukturen entwickeln sich in Stadien, von blassen, leeren Hüllen zu dunklen, mit Melanin gefüllten Granula, die zur Zelloberfläche transportiert werden können. Ihre Aktivität wird durch Signale wie das Hormon α‑MSH gesteuert, das nach UV‑Licht ansteigt und die Pigmentierung fördert. Damit Melanosomen richtig funktionieren, müssen sich ihre inneren chemischen Verhältnisse — insbesondere Säuregrad (pH) und Calciumspiegel — zur richtigen Zeit verändern. Ein früher Schritt ist die Bildung eines Proteingerüsts aus PMEL‑Fibrillen, das ein saures Innenmilieu erfordert. Später wird das Kompartiment weniger sauer, damit melaninproduzierende Enzyme arbeiten können. Wie diese präzisen Änderungen angetrieben und zeitlich gesteuert werden, war bisher unklar.
Wenn Kraftwerke auf Pigmentgranula treffen Figure 1.
Das Team konzentrierte sich auf physische Kontaktstellen zwischen Mitochondrien und Melanosomen. Solche Organellen‑„Handschläge“ sind an anderen Stellen in Zellen bereits als wichtig bekannt, etwa zwischen Mitochondrien und dem endoplasmatischen Retikulum. Hier entwickelten die Forschenden ein Lebendzell‑Reporter‑System namens MiMSBiT, das leuchtet, wenn Mitochondrien und Melanosomen nahe genug zusammenkommen, damit sich zwei gentechnisch veränderte Proteinfragmente wieder vereinen. Mit diesem Werkzeug in Maus‑Melanomzellen fanden sie, dass α‑MSH und verwandte Signale einen starken, aber vorübergehenden Anstieg der Mitochondrien–Melanosomen‑Kontakte bewirkten. Diese Kontakte erreichten etwa drei Stunden nach Stimulation ihr Maximum — das gleiche Zeitfenster, in dem Melanosomen am sauersten wurden und PMEL‑Fibrillen gebildet wurden — was darauf hindeutet, dass die physische Nähe der beiden Organellen eng mit der Reifung der Pigmentgranula verknüpft ist.
Das Verbindungs‑Team: STIM1 und Mitofusin 2
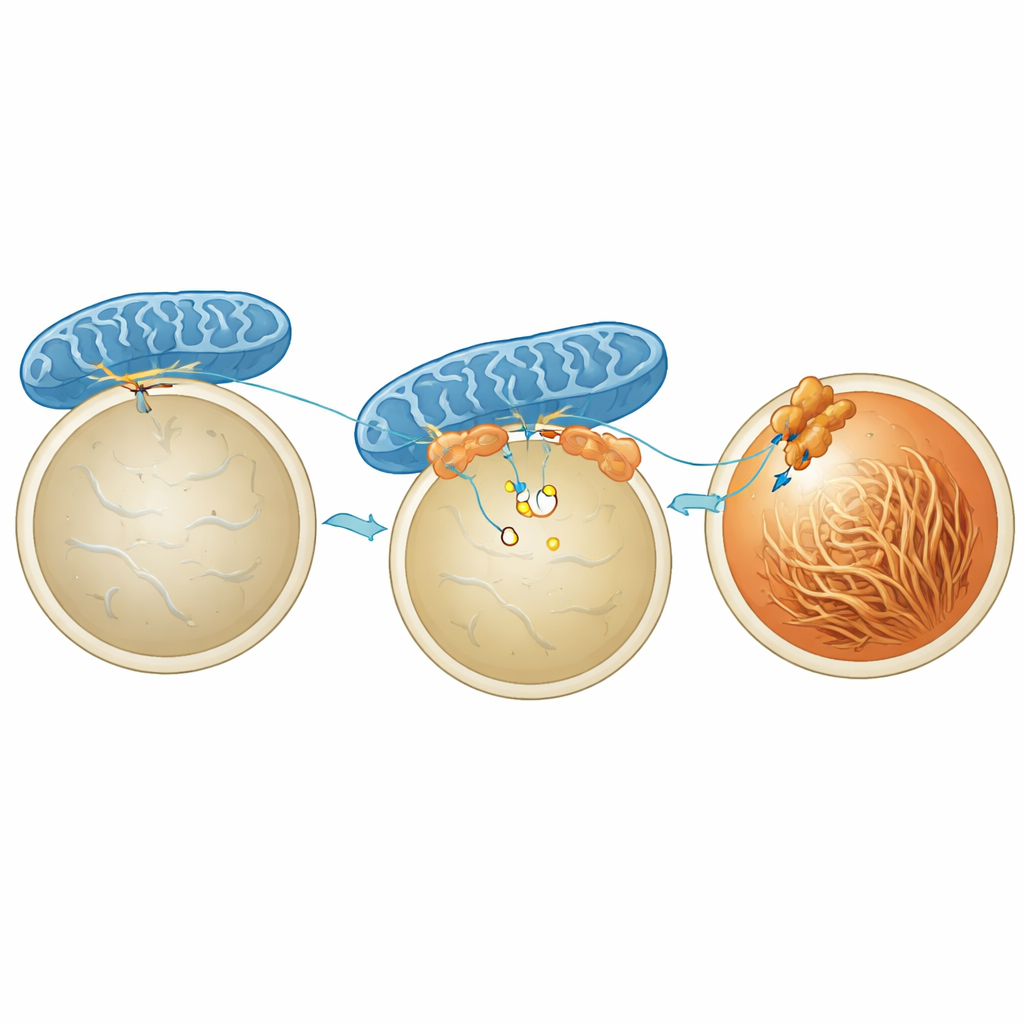
Um zu verstehen, was Mitochondrien und Melanosomen tatsächlich zusammenhält, fokussierten sich die Wissenschaftler auf ein Protein namens MFN2, das bereits dafür bekannt ist, Mitochondrien mit anderen Organellen zu verbinden. Das Herunterregulieren von MFN2 in Pigmentzellen verringerte die hormoninduzierten Kontakte stark und schwächte die Zunahme der Pigmentierung ab, ohne die Mengen oder die Grundaktivität der melaninproduzierenden Enzyme zu verändern. Der entscheidende Akteur auf der Seite des Melanosoms erwies sich als STIM1, besser bekannt als Calciumsensor in einem anderen Zellkompartiment. Mithilfe einer Proximity‑Labeling‑Methode und hochauflösender Bildgebung zeigten die Forschenden, dass ein Pool von STIM1 auf Melanosomen sitzt und bei Anwesenheit von α‑MSH kurzzeitig an MFN2 auf Mitochondrien bindet. Diese Interaktion wird durch einen kurzlebigen Calciumabfall im Lumen des Melanosoms ausgelöst, der dazu führt, dass STIM1 sich klumpt und MFN2 greift, wodurch eine physische Brücke entsteht.
Energiezufuhr und Versauerung innerhalb von Melanosomen Figure 2.
Was gewinnt man dadurch, Mitochondrien und Melanosomen zusammenzuziehen? Die Studie zeigt, dass diese Kontakte lokal die Verfügbarkeit von ATP, der Energiewährung der Zelle, direkt an der Melanosomoberfläche erhöhen. Mit einem fluoreszenten ATP‑Sensor, der an der Melanosomenmembran verankert ist, fanden die Autorinnen und Autoren heraus, dass α‑MSH das ATP um Melanosomen erhöht — auf eine Weise, die von mitochondrialer Energieproduktion abhängt, aber nicht vom Zuckerabbau im Zytoplasma. Wenn MFN2 oder STIM1 reduziert wurden, verschwand dieser lokale ATP‑Anstieg, obwohl die Gesamtzahl der Kontakte oder der grobe Zellstoffwechsel nicht dramatisch verändert war. Das zusätzliche ATP scheint Protonenpumpen in der Melanosomenmembran anzutreiben, die aktiv Protonen einpumpen und das Innenleben vorübergehend ansäuern. Dieser Säureimpuls fördert wiederum die Assemblierung von PMEL zu geordneten Fibrillen, die als Gerüst dienen, auf dem später sicher Melanin abgelagert werden kann.
Von Zellkontakten zur Pigmentierung des ganzen Körpers
Um zu testen, ob dieser mikroskopische Mechanismus auch in lebenden Tieren relevant ist, behandelten die Forschenden Zebrafisch‑Embryonen mit einem Wirkstoff, der die gleiche MFN2‑Region stört, die für die STIM1‑Bindung benötigt wird. Die sich entwickelnden Fische zeigten deutlich blassere Körper, was bestätigt, dass die Störung dieser Organellenkontakte die normale Pigmentierung in vivo beeinträchtigt. Zusammen zeichnen die Ergebnisse eine schrittweise Geschichte nach: Hormonelle Signale verursachen Calciumveränderungen innerhalb unreifer Melanosomen; das aktiviert STIM1, das sich mit MFN2 zusammenschließt, um Mitochondrien zu fixieren; diese Mitochondrien liefern dann ATP genau dorthin, wo es gebraucht wird, um Melanosomen zu ansäuern und das PMEL‑Gerüst zu organisieren; und erst dann kann eine robuste Melaninproduktion beginnen. Für eine Laienbetrachterin bedeutet das: Die Farbe, die wir auf Haut und Haaren sehen, hängt von exquisit getimten, nanoskaligen Wechselwirkungen zwischen winzigen Strukturen tief in jeder Pigmentzelle ab.
Zitation: Shiiba, I., Ishikawa, Y., Oshio, H. et al. STIM1-Mitofusin2 interactions tether mitochondria and melanosome contacts that promote melanosome maturation.
Nat Commun17, 3593 (2026). https://doi.org/10.1038/s41467-026-70282-w
Schlüsselwörter: Mitochondrien–Melanosomen-Kontakt, Melaninproduktion, Organelkommunikation, Hautpigmentierung, zelluläre Energie und ATP