Clear Sky Science · pl
Fosforylacyjnie zależny przełącznik ubikwitynacji orkiestruje jądrowe przeprogramowanie odporności po rozpoznaniu chityny
Jak ryż broni się przed wyniszczającą chorobą upraw
Rolnicy ryżu na całym świecie zmagają się z chorobą pleśniową (blast), grzybiczym zakażeniem, które może zniszczyć pola i zagrażać bezpieczeństwu żywnościowemu. W tym badaniu odkryto, jak rośliny ryżu wykorzystują drobny fragment ściany komórkowej grzyba, zwany chityną, jako sygnał alarmowy, a następnie przełączają wewnętrzny mechanizm molekularny, by uruchomić silne mechanizmy obronne w jądrze komórkowym. Zrozumienie tego przełącznika nie tylko wyjaśnia długo brakujący etap w odporności roślin, lecz może też ukierunkować hodowlę lub inżynierię odporniejszych odmian ryżu.
Powierzchniowy alarm dociera głęboko do wnętrza komórki
Rośliny nie mogą uciekać przed infekcją, dlatego polegają na wbudowanych czujnikach na powierzchni komórek. W ryżu jeden z takich receptorów wykrywa chitynę, kluczowy składnik osłonki grzyba powodującego blast. Wcześniejsze prace wykazały, że po wykryciu chityny na powierzchni komórki enzym pomocniczy o nazwie OsRLCK185 przekazuje sygnał i pomaga wywołać wczesne reakcje obronne, takie jak wyrzuty reaktywnych form tlenu. Nadal tajemnicze było to, jak ten wczesny sygnał dociera do jądra, gdzie włączane lub wyłączane są geny kontrolujące długotrwałą odporność. Artykuł wypełnia tę lukę, śledząc bezpośrednią drogę od sensora chityny przez OsRLCK185 do kluczowych regulatorów przekształcających aktywność genów w jądrze.
Pomocni strażnicy i destrukcyjna etykieta
Autorzy koncentrują się na dwóch blisko spokrewnionych białkach, OsGF14f i OsGF14c, należących do rodziny 14-3-3, które często pełnią rolę molekularnych „eskort” lub stabilizatorów. Poprzez wyłączenie genów lub nadprodukcję tych białek w ryżu wykazali, że zarówno OsGF14f, jak i OsGF14c zwiększają odporność roślin na grzyba blast: rośliny pozbawione ich mają większe ogniska choroby i intensywniejszy rozwój grzyba, podczas gdy rośliny z dodatkowymi kopiami są lepiej chronione. Jednak w komórce istnieje też wbudowany hamulec: enzym nazwany OsPUB20, który przyłącza małe znaczniki molekularne (ubikwitynę) do OsGF14f i OsGF14c. To znakowanie kieruje je do zniszczenia przez komórkową maszynerię do rozkładu białek, osłabiając obronę rośliny. Pokrewny enzym, OsPUB19, odgrywa podobną negatywną rolę, a rośliny pozbawione obu enzymów stają się szczególnie odporne, ponieważ ich ochronne białka 14-3-3 akumulują się.
Przełącznik fosforylacyjny, który oszczędza kluczowego obrońcę
Percepcja chityny robi więcej niż tylko uruchamia alarm; zmienia też działanie OsPUB20. Zespół wykazał, że OsRLCK185 fizycznie wiąże się z OsPUB20 i dodaje małą grupę fosforanową w jednym miejscu (reszcie aminokwasowej treoniny 153). Ta zmiana chemiczna nie wyłącza OsPUB20 całkowicie, lecz osłabia jego oddziaływanie specyficznie z OsGF14f. W rezultacie OsGF14f jest mniej intensywnie znakowany ubikwityną i unika degradacji, podczas gdy OsGF14c pozostaje bardziej podatne. Rośliny zaprojektowane tak, aby miały wariant OsPUB20 imitujący stałą fosforylację, chronią OsGF14f i wykazują silniejszą odporność na blast, natomiast rośliny z niefosforylowalną wersją tracą więcej OsGF14f i stają się bardziej wrażliwe. W ten sposób OsRLCK185 działa jak precyzyjny przełącznik, który selektywnie zabezpiecza jednego z najważniejszych obrońców rośliny.
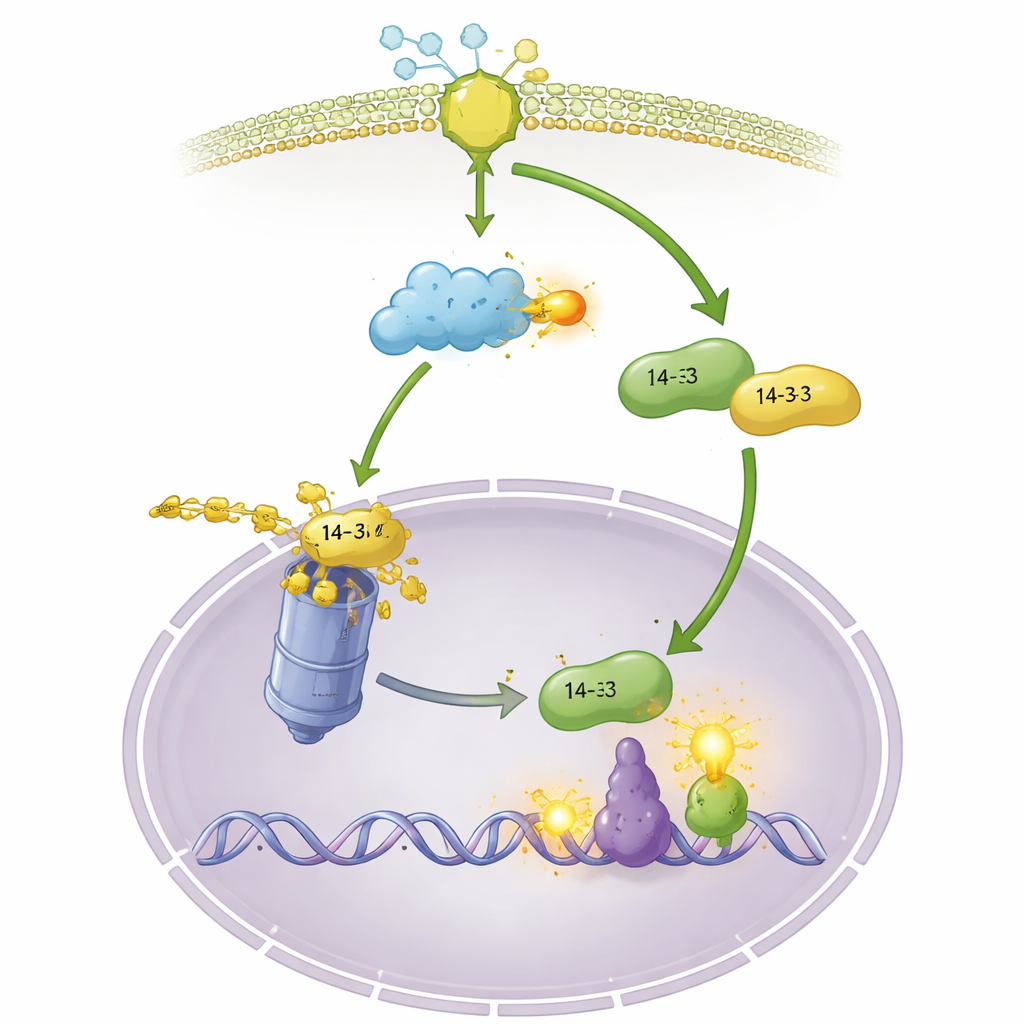
Z płynu komórkowego do centrum sterowania
Stabilizacja OsGF14f to tylko część historii. Autorzy odkryli, że po traktowaniu chityną lub po infekcji grzybem OsGF14f przemieszcza się z płynu komórkowego do jądra, podczas gdy OsGF14c w większości pozostaje na zewnątrz. Wymuszenie lokalizacji OsGF14f w jądrze sprawia, że rośliny są jeszcze bardziej odporne, co podkreśla, jak ważna jest ta relokacja. Wewnątrz jądra OsGF14f wiąże się z czynnikiem transkrypcyjnym OsWRKY42, który normalnie działa jako hamulec obrony, tłumiąc geny związane z hormonami. OsGF14f sprzyja rozkładowi OsWRKY42 za pośrednictwem tej samej maszynerii degradacji białek, usuwając ten hamulec. Rośliny pozbawione OsWRKY42 są bardziej odporne, a rośliny bogate w OsGF14f wykazują wyższą aktywność genów obronnych związanych zarówno z kwasem jasmonowym, jak i kwasem salicylowym — dwoma kluczowymi hormonami odpornościowymi. Tak więc OsGF14f pełni rolę przekaźnika łączącego cytoplazmatyczny alarm z jądrowym przeprogramowaniem genów obronnych.
Dlaczego ten obwód molekularny ma znaczenie
Podsumowując, praca ujawnia wielowarstwowy układ obronny w ryżu. Gdy chityna z grzyba blast jest wykrywana na powierzchni komórki, OsRLCK185 modyfikuje OsPUB20, osłabiając jego zdolność do niszczenia OsGF14f. OsGF14f, które uniknęło degradacji, następnie kumuluje się i przemieszcza do jądra, gdzie pomaga usunąć represor OsWRKY42 i uwalnia ważne szlaki obronne do działania. Ten zależny od fosforylacji „przełącznik ubikwitynacji” elegancko łączy zewnętrzny sygnał zagrożenia z głębokimi zmianami aktywności genów, pomagając roślinie przełączyć się z normalnego wzrostu na zwiększoną ochronę. Mapując tę ścieżkę, badanie wskazuje obiecujące cele do programów hodowlanych lub inżynieryjnych, które mogą umożliwić tworzenie odmian ryżu lepiej odpornych na blast przy mniejszym użyciu chemikaliów.
Cytowanie: Zhang, C., Suttiviriya, P., Wang, R. et al. A phosphorylation-dependent ubiquitination switch orchestrates nuclear immune reprogramming upon chitin perception. Nat Commun 17, 2998 (2026). https://doi.org/10.1038/s41467-026-69627-2
Słowa kluczowe: choroba pleśni ryżu, odporność roślin, degradacja białek, transdukcja sygnału, ochrona upraw