Clear Sky Science · de
Ein phosphorylationsabhängiger Ubiquitinierungsschalter orchestriert nukleare Immun‑Reprogrammierung nach Chitinwahrnehmung
Wie Reis einer verheerenden Kulturkrankheit trotzt
Reisbauern weltweit kämpfen gegen die Brandkrankheit, eine Pilzinfektion, die Felder vernichten und die Ernährungssicherheit bedrohen kann. Diese Studie zeigt, wie Reis Pflanzen ein winziges Fragment der Pilzzellwand, das Chitin, als Alarmsignal nutzt und anschließend einen inneren molekularen Schalter umlegt, um starke Abwehrreaktionen im Zellkern zu aktivieren. Das Verständnis dieses Schalters erklärt nicht nur einen lange fehlenden Schritt der Pflanzenimmunität, sondern kann auch Züchtung oder biotechnologische Verbesserungen robusterer Reissorten leiten.
Ein Oberflächenalarm dringt tief in die Zelle vor
Pflanzen können einer Infektion nicht ausweichen, deshalb verlassen sie sich auf eingebaute Sensoren an der Zelloberfläche. Beim Reis erkennt einer dieser Sensoren Chitin, einen zentralen Bestandteil der Hülle des Brandpilzes. Frühere Arbeiten zeigten, dass nach der Chitinwahrnehmung an der Zelloberfläche ein Helferenzym namens OsRLCK185 das Signal weiterleitet und frühe Abwehrreaktionen wie reaktive Sauerstoff‑S töße auslöst. Unklar blieb jedoch, wie dieses frühe Signal den Nukleus erreicht, wo Gene, die die langfristige Immunität steuern, an- oder abgeschaltet werden. Diese Arbeit schließt diese Lücke, indem sie einen direkten Weg vom Chitin‑Sensor über OsRLCK185 zu Schlüsselregulatoren nachzeichnet, die die genaktiveit im Zellkern umgestalten.
Hilfreiche Wächter und ein zerstörerisches Etikett
Die Autoren konzentrieren sich auf zwei eng verwandte Proteine, OsGF14f und OsGF14c, Mitglieder der 14‑3‑3‑Familie, die häufig als molekulare „Begleiter“ oder Stabilisatoren fungieren. Durch das Ausschalten oder die Überexpression dieser Proteine im Reis zeigen sie, dass sowohl OsGF14f als auch OsGF14c die Pflanzen gegen den Brandpilz widerstandsfähiger machen: Pflanzen ohne diese Proteine entwickeln größere Krankheitsflecken und stärkeren Pilzwuchs, während Pflanzen mit zusätzlichen Kopien besser geschützt sind. Die Zelle besitzt jedoch eine eingebaute Bremse: ein Enzym namens OsPUB20, das kleine molekulare Anhängsel (Ubiquitin) an OsGF14f und OsGF14c heftet. Diese Markierung signalisiert ihren Abbau durch die zelluläre Protein‑Vernichtungsmaschinerie und schwächt so die Abwehr. Ein verwandtes Enzym, OsPUB19, spielt eine ähnliche negative Rolle, und Pflanzen, denen beide Enzyme fehlen, sind besonders widerstandsfähig, weil sich die schützenden 14‑3‑3‑Proteine ansammeln.
Ein Phosphorylierungsschalter, der einen Schlüsselverteidiger verschont
Die Chitinwahrnehmung bewirkt mehr als nur das Auslösen eines Alarms; sie verändert das Verhalten von OsPUB20. Das Team zeigt, dass OsRLCK185 physisch an OsPUB20 bindet und an einer einzigen Position (der Aminosäure Threonin 153) eine Phosphatgruppe anhängt. Diese chemische Modifikation schaltet OsPUB20 nicht vollständig ab, schwächt aber seine Bindung speziell an OsGF14f. Infolgedessen wird OsGF14f weniger stark mit Ubiquitin markiert und entgeht der Zerstörung, während OsGF14c weiterhin anfälliger bleibt. Pflanzen, die so verändert wurden, dass OsPUB20 dauerhaft phosphoryliert erscheint, schützen OsGF14f und zeigen stärkere Brandresistenz, während Pflanzen mit einer nicht phosphorylierbaren Variante mehr OsGF14f verlieren und anfälliger werden. Auf diese Weise wirkt OsRLCK185 als ein feineingestellter Schalter, der selektiv einen der wichtigsten Verteidiger der Pflanze bewahrt.
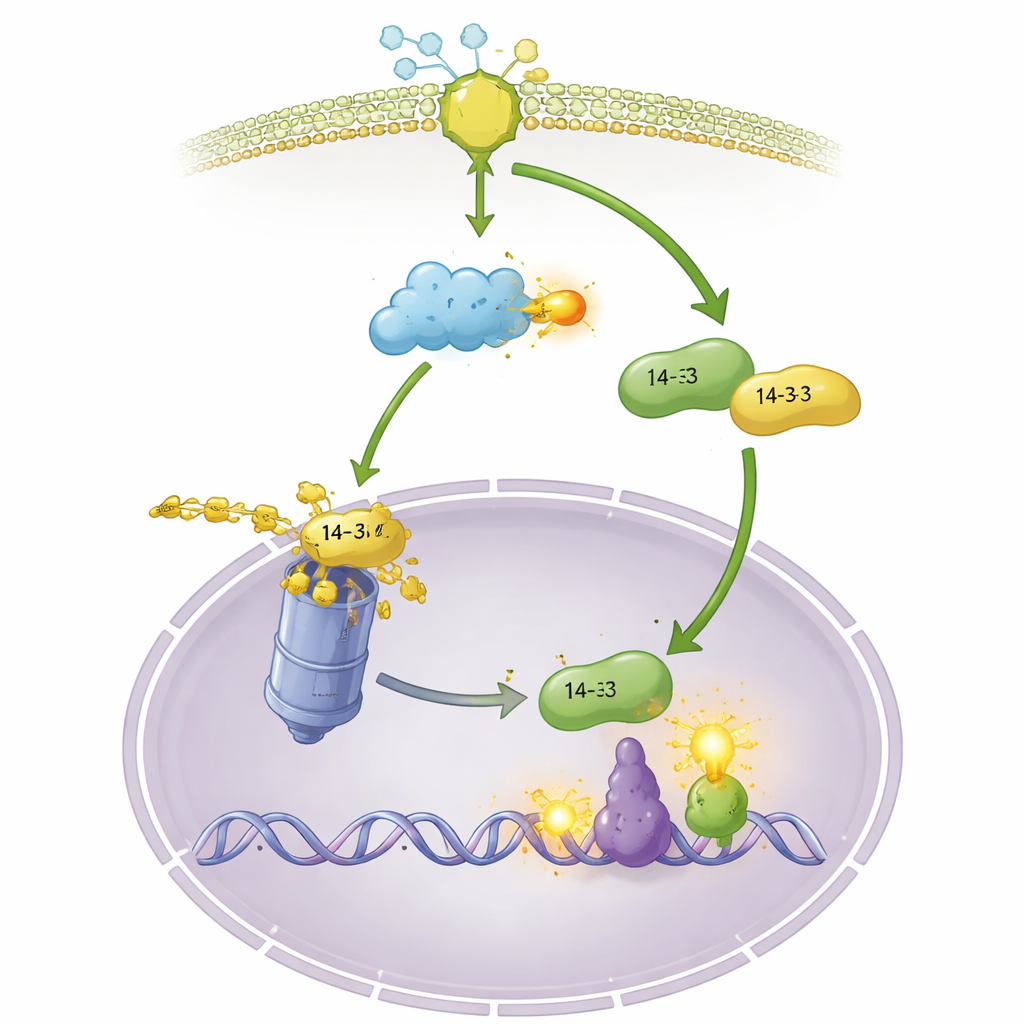
Vom Zellplasma ins Kontrollzentrum
Die Stabilisierung von OsGF14f ist nur ein Teil der Geschichte. Die Autoren stellen fest, dass sich nach Chitinbehandlung oder Pilzinfektion OsGF14f vom Zellplasma in den Nukleus bewegt, während OsGF14c überwiegend außerhalb bleibt. Das Erzwingen der Kernlokalisierung von OsGF14f macht die Pflanzen noch widerstandsfähiger und unterstreicht, wie entscheidend diese Umsiedlung ist. Im Nukleus bindet OsGF14f an einen Transkriptionsfaktor namens OsWRKY42, der normalerweise als Bremse wirkt, indem er hormonbezogene Gene dämpft. OsGF14f fördert den Abbau von OsWRKY42 über dieselbe Proteinabbau‑Maschinerie und löst damit diese Bremse. Pflanzen ohne OsWRKY42 sind widerstandsfähiger, und Pflanzen mit hohem OsGF14f‑Ge halt zeigen eine stärkere Aktivität von Abwehrgenen, die mit Jasmon‑ und Salicylsäure verbunden sind, zwei wichtigen Immunhormonen. So dient OsGF14f als Shuttle, das den cytoplasmatischen Alarm mit der nuklearen Reprogrammierung von Abwehrgenen verbindet.
Warum dieser molekulare Schaltkreis wichtig ist
Zusammengefasst enthüllt die Arbeit einen geschichteten Verteidigungszyklus im Reis. Wenn Chitin vom Brandpilz an der Zelloberfläche erkannt wird, modifiziert OsRLCK185 OsPUB20 und schwächt dessen Fähigkeit, OsGF14f zu zerstören. Das verschonte OsGF14f sammelt sich an und wandert in den Nukleus, wo es hilft, den Repressor OsWRKY42 zu entfernen und wichtige Abwehrwege zu aktivieren. Dieser phosphorylationsabhängige „Ubiquitinierungs‑Schalter“ verbindet auf elegante Weise ein äußerliches Gefahrenignal mit tiefgreifenden Veränderungen der Genaktivität und hilft der Pflanze, vom normalen Wachstum in einen Zustand erhöhter Verteidigung umzuschalten. Durch die Kartierung dieses Weges hebt die Studie vielversprechende Ziele für die Züchtung oder das Engineering von Reissorten hervor, die der Brandkrankheit besser standhalten und weniger Chemikalien benötigen.
Zitation: Zhang, C., Suttiviriya, P., Wang, R. et al. A phosphorylation-dependent ubiquitination switch orchestrates nuclear immune reprogramming upon chitin perception. Nat Commun 17, 2998 (2026). https://doi.org/10.1038/s41467-026-69627-2
Schlüsselwörter: Reisbrandkrankheit, Pflanzenimmunität, Proteinabba u, Signaltransduktion, Pflanzenschutz