Clear Sky Science · pl
Caspaza-3/Drice jako kluczowy regulator dynamiki aktyny poprzez podwójną kontrolę małej rodziny RhoGTPaz i Gelsoliny w cewkach Malpighiego Drosophila
Dlaczego rusztowania komórkowe są ważne dla małych nerek
Każda komórka w naszym organizmie polega na mikroskopijnym rusztowaniu zbudowanym z filamentów aktynowych, aby utrzymać kształt, poruszać się i tworzyć tkanki. Niniejsze badanie analizuje, jak powszechnie znany „enzym śmierci” u muszek owocowych pełni nieoczekiwaną podwójną rolę — jako budowniczy, który utrzymuje to rusztowanie w równowadze, tak by ich rurkowate, przypominające nerki struktury prawidłowo rosły i funkcjonowały. Ponieważ podobne systemy działają u wyższych zwierząt, w tym u ludzi, praca ta wskazuje nowe sposoby, w jakie enzymy związane ze śmiercią komórkową mogą wpływać na zdrowy rozwój i chorobę.
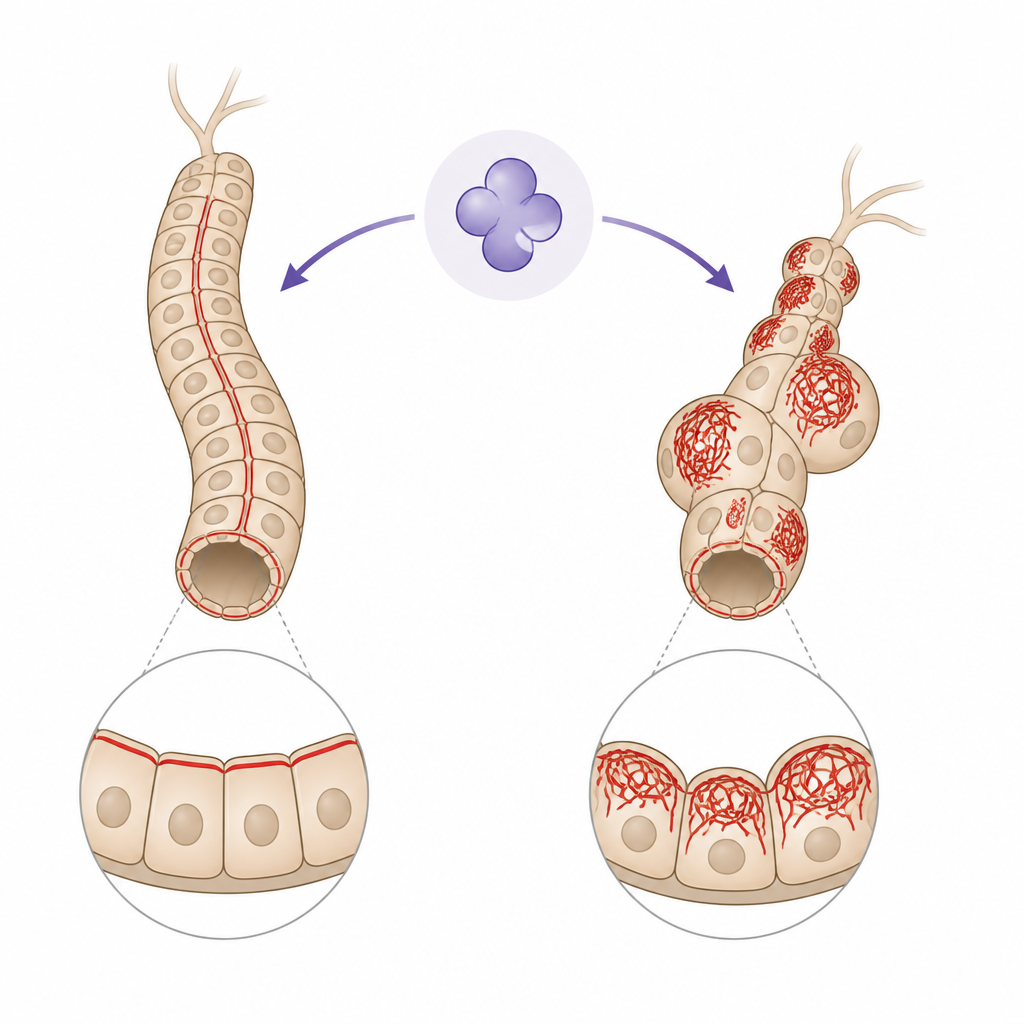
Zaskakująca rola enzymu śmierci
Caspazy zwykle przedstawiane są jako molekularni wykonawcy demontujący komórki podczas zaprogramowanej śmierci komórkowej. Autorzy koncentrują się na Drice, muszkowym odpowiedniku caspazy 3, w cewkach Malpighiego — wąskich rurkach pełniących funkcję nerek owadów. Te rurki działają przez istotne etapy życia bez ulegania zniszczeniu, mimo obecności aktywnego Drice. Wcześniejsze prace wykazały, że gdy Drice brakuje, rurki stają się krótsze, wypełnione pęcherzami z płynem i naszpikowane nadmiernie gęstymi filamentami aktyny. Nowe badanie pyta, jak Drice zapobiega temu strukturalnemu chaosowi i które ścieżki sygnałowe łączą go z siecią aktynową.
Utrzymywanie porządku w nitkach aktyny
Aktyna nieustannie przełącza się między wolnymi monomerami a zmontowanymi filamentami, a tę równowagę kontroluje wiele białek pomocniczych. Jednym z kluczowych węzłów jest rodzina molekularnych przełączników Rho, które regulują gdzie i kiedy powstaje aktyna. W normalnych cewkach jeden z tych przełączników, Rho1, aktywuje partnera o nazwie Rok, pomagając zorganizować cienkie, regularnie ułożone filamenty aktyny na powierzchni komórki. U muszek-mutantów pozbawionych Drice poziomy Roka spadają, filamenty tracą uporządkowanie, a polaryzacja tkanki od wierzchu do dołu ulega zaburzeniu. Gdy badacze przywrócili Roka w tle mutanta, zarówno organizacja aktyny, jak i kształt rurki poprawiły się, co pokazuje, że Drice pomaga utrzymać uporządkowane rusztowania częściowo przez wspieranie szlaku Rho1–Rok.
Kiedy sygnały wzrostu stają się zbyt intensywne
Zespół przebadał następnie innego członka rodziny Rho, Cdc42, który normalnie sprzyja rozgałęzianiu filamentów aktynowych przez kompleks partnerów Arp2/3. W cewkach pozbawionych Drice poziomy Cdc42 i obu komponentów Arp były zwiększone. Ten wzrost prowadził do grubszego, silniej rozgałęzionego aktynu wzdłuż kory komórkowej. Stłumienie Arp2 lub Arp3 spowodowało przerzedzenie filamentów i ich zbliżenie do wyglądu z zdrowych cewek oraz częściowe przywrócenie ogólnego uporządkowania nawet u mutantów Drice. Wyniki sugerują, że bez Drice jedna z gałęzi systemu promujących rozrost rozgałęzień aktyny staje się nadaktywna, przyczyniając się do gęstej, splątanej sieci deformującej kształt komórek.
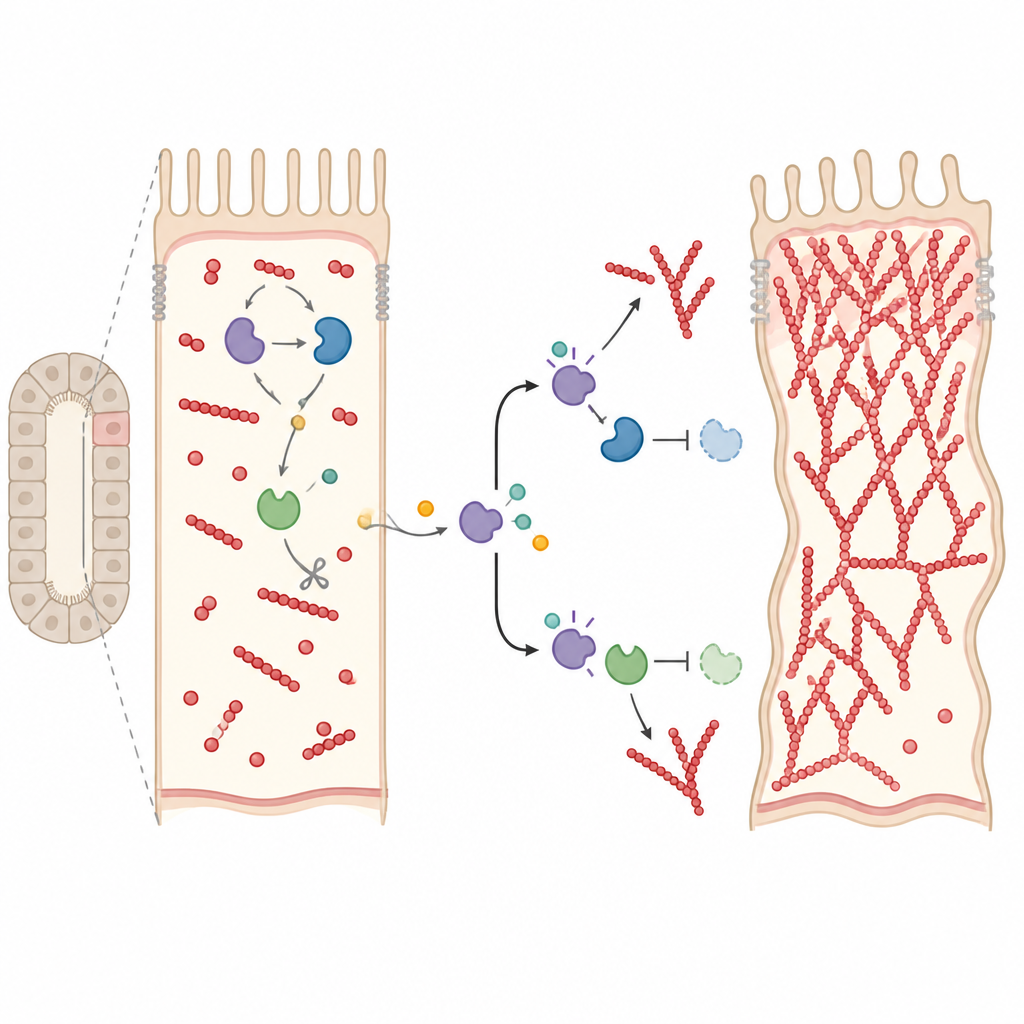
Zepsany hamulec wzrostu filamentów
Aby zrozumieć, jak Drice łączy się z tymi przełącznikami, autorzy poszukiwali białek fizycznie wchodzących w interakcje z Rho1 w normalnych i mutantnych cewkach. Stwierdzili, że Gelsolina — białko, które normalnie przecina i nasadawia końce filamentów aktynowych, aby zatrzymać dalszy wzrost — łączy się z Rho1 tylko obecności Drice. Drice również aktywuje Gelsolinę przez jej przecięcie, krok znacząco zmniejszony u mutantów. W cewkach z wyciszoną ekspresją Drice i Gelsoliny stosunek zmontowanych filamentów do wolnej aktyny przesuwał się zdecydowanie w stronę filamentów, wskazując na niekontrolowaną polimeryzację. Co znamienne, podbicie poziomu Roka u mutantów Drice przywróciło ten stosunek blisko normy, sugerując że Drice koordynuje zarówno sygnały „startu”, jak i „stopu” wzrostu aktyny przez ścieżki Rok, Cdc42–Arp2/3 oraz Gelsolinę.
Jak to zmienia nasze spojrzenie na białka śmierci komórkowej
Śledząc zmiany w sygnalizacji, interakcjach białkowych i strukturze aktyny, badanie pokazuje, że Drice nie jest jedynie niszczycielem, lecz także opiekunem architektury komórkowej w przypominających nerki cewkach muszek owocowych. Gdy Drice jest nieobecny, szlaki promujące wzrost wymykają się spod kontroli, hamulce długości filamentów zawodzą, a delikatna równowaga między wolną a zmontowaną aktyną zostaje utracona, prowadząc do zdeformowanych, źle funkcjonujących rurek. Dla czytelnika popularnonaukowego kluczowe przesłanie brzmi: enzymy znane z wywoływania śmierci komórkowej mogą również odgrywać zasadniczą rolę w budowaniu i utrzymaniu żywych tkanek, a podobne systemy regulacyjne mogą wpływać na rozwój, naprawę i reakcje naszych własnych narządów na stres.
Cytowanie: Sagar, S.C., Tapadia, M.G. Caspase-3/Drice as a critical regulator of actin dynamics through its dual control of small RhoGTPase family and Gelsolin in the Malpighian tubules of Drosophila. Cell Death Discov. 12, 214 (2026). https://doi.org/10.1038/s41420-026-03061-7
Słowa kluczowe: cytoszkielet aktynowy, caspaza-3, Rho GTPazy, Gelsolina, cewki Malpighiego Drosophila