Clear Sky Science · fr
Caspase-3/Drice en tant que régulateur critique de la dynamique de l’actine via son double contrôle des petites RhoGTPases et de la Gelsoline dans les tubules de Malpighi de Drosophila
Pourquoi les échafaudages cellulaires comptent pour les petits reins
Chaque cellule de notre corps dépend d’un échafaudage microscopique composé de filaments d’actine pour conserver sa forme, se déplacer et construire des organes. Cette étude examine comment une « enzyme de la mort » bien connue chez la mouche des fruits joue, de façon inattendue, un rôle de bâtisseuse, en maintenant cet échafaudage en équilibre afin que leurs tubules de type rénal se développent et fonctionnent correctement. Comme des systèmes similaires existent chez les animaux supérieurs, y compris les humains, ce travail suggère de nouvelles façons dont les enzymes impliquées dans la mort cellulaire pourraient influencer le développement sain et les maladies.
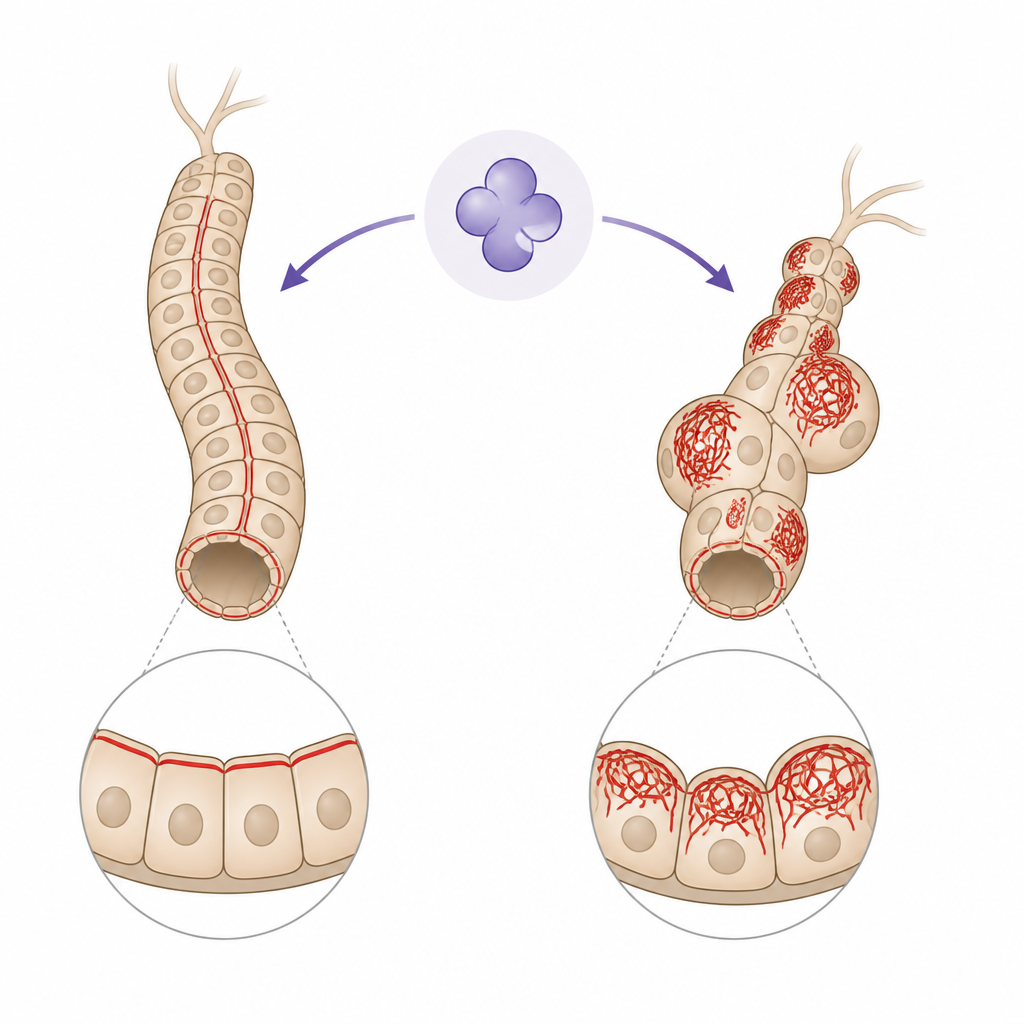
Un rôle surprenant pour une enzyme de la mort
Les caspases sont généralement présentées comme des exécuteurs moléculaires qui démantèlent les cellules lors de la mort cellulaire programmée. Les auteurs se concentrent sur Drice, la version de la caspase 3 chez la mouche, dans les tubules de Malpighi, un ensemble de tubes étroits qui font office de reins chez l’insecte. Ces tubules continuent de fonctionner à travers d’importants changements de vie sans être détruits, bien qu’ils contiennent Drice actif. Des travaux antérieurs ont montré que, lorsque Drice fait défaut, les tubules deviennent plus courts, couverts de kystes remplis de liquide et encombrés de filaments d’actine excessivement denses. La nouvelle étude s’interroge sur la façon dont Drice empêche ce chaos structurel et quels chemins de signalisation le relient au réseau d’actine.
Maintenir l’ordre des brins d’actine
L’actine passe en permanence d’unités libres à des filaments assemblés, et cet équilibre est contrôlé par de nombreuses protéines auxiliaires. Un centre clé est la famille Rho de commutateurs moléculaires, qui ajustent où et quand l’actine se forme. Dans des tubules normaux, l’un de ces commutateurs, Rho1, active un partenaire appelé Rok pour aider à organiser des filaments d’actine fins et régulièrement alignés le long de la surface cellulaire. Chez les mouches mutantes pour Drice, les niveaux de Rok chutent, les filaments d’actine perdent leur arrangement ordonné et la polarité du tissu de haut en bas se rompt. Lorsque les chercheurs ont restauré Rok dans le contexte mutant, à la fois l’organisation de l’actine et la forme du tube se sont améliorées, montrant que Drice contribue à maintenir des échafaudages ordonnés en soutenant en partie la voie Rho1–Rok.
Quand les signaux de croissance s’emballent
L’équipe a ensuite examiné un autre membre de la famille Rho, Cdc42, qui favorise normalement la ramification des filaments d’actine via un complexe partenaire appelé Arp2/3. Dans les tubules dépourvus de Drice, Cdc42 et les deux composants d’Arp ont été augmentés. Cette poussée a conduit à une actine plus épaisse et plus fortement branchée le long du cortex cellulaire. Réduire l’expression d’Arp2 ou d’Arp3 a rendu les filaments plus fins et plus similaires à ceux des tubules sains, et a partiellement restauré l’alignement global même chez les mutants Drice. Ces résultats suggèrent que sans Drice, une branche du système qui favorise la croissance des ramifications d’actine devient hyperactive, contribuant à un réseau dense et emmêlé qui déforme la forme cellulaire.
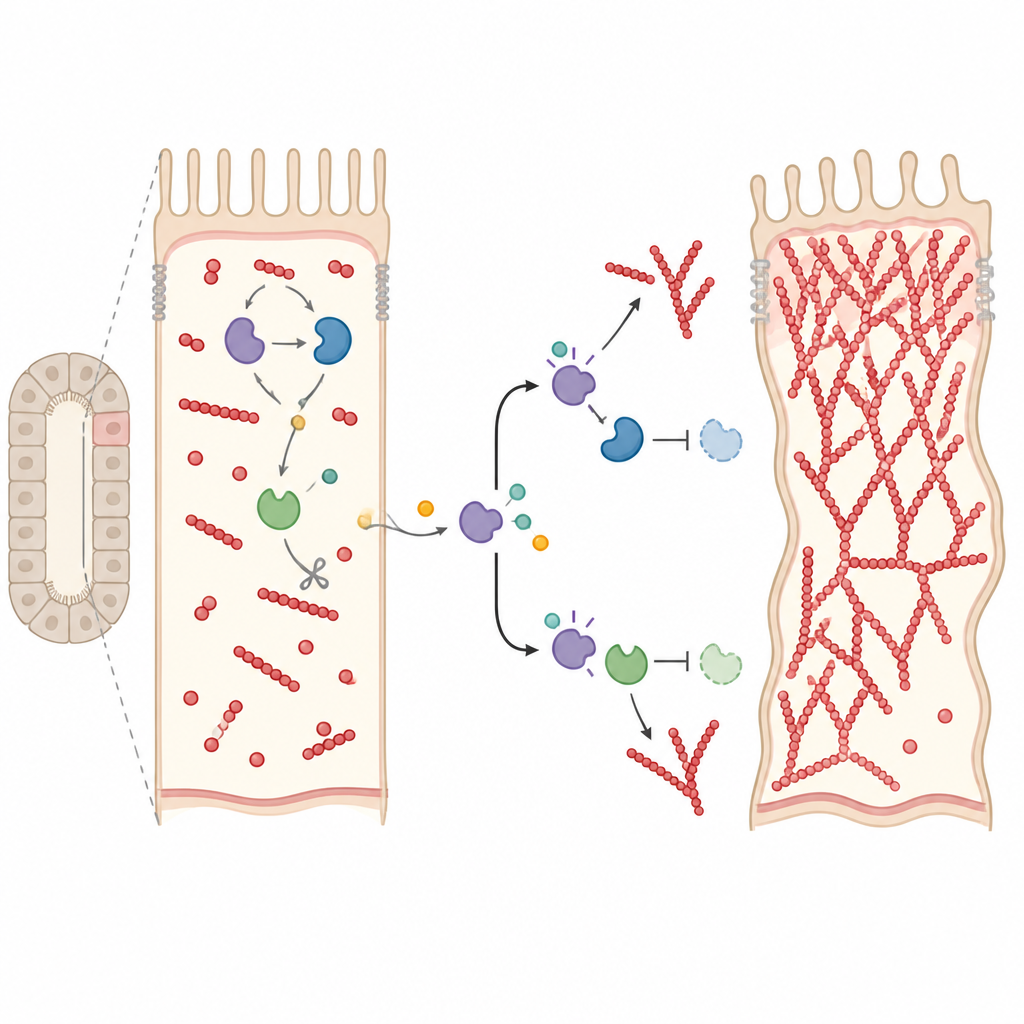
Un frein cassé sur la croissance des filaments
Pour comprendre comment Drice s’intègre à ces commutateurs, les auteurs ont recherché des protéines interagissant physiquement avec Rho1 dans des tubules normaux et mutants. Ils ont trouvé que la Gelsoline, une protéine qui coupe et coiffe normalement les filaments d’actine pour arrêter leur croissance, s’associe à Rho1 seulement lorsque Drice est présent. Drice active aussi la Gelsoline en la clivant, une étape fortement réduite chez les mutants. Dans les tubules où Drice ou la Gelsoline sont réduits, le ratio filaments assemblés/actine libre bascule nettement en faveur des filaments, indiquant une polymérisation incontrôlée. Remarquablement, renforcer Rok chez les mutants Drice a ramené ce ratio proche de la normale, suggérant que Drice coordonne à la fois les signaux « go » et « stop » sur la croissance de l’actine via Rok, Cdc42–Arp2/3 et la Gelsoline.
Comment cela change notre vision des protéines de la mort cellulaire
En suivant les changements de signalisation, d’interactions protéiques et de structure de l’actine, l’étude montre que Drice n’est pas seulement un destructeur mais aussi un gardien de l’architecture cellulaire dans les tubules de type rénal chez la mouche. Lorsqu’il manque, les voies favorisant la croissance dépassent leur cible, les freins sur la longueur des filaments échouent et l’équilibre fin entre actine libre et assemblée est perdu, conduisant à des tubes déformés et mal fonctionnants. Pour le lecteur non spécialiste, le message clé est que des enzymes célèbres pour tuer les cellules peuvent aussi être essentielles à la construction et au maintien des tissus vivants, et que des systèmes de contrôle similaires peuvent influencer la façon dont nos propres organes se développent, se réparent et réagissent au stress.
Citation: Sagar, S.C., Tapadia, M.G. Caspase-3/Drice as a critical regulator of actin dynamics through its dual control of small RhoGTPase family and Gelsolin in the Malpighian tubules of Drosophila. Cell Death Discov. 12, 214 (2026). https://doi.org/10.1038/s41420-026-03061-7
Mots-clés: cytosquelette d’actine, caspase-3, Rho GTPases, Gelsoline, tubules de Malpighi de Drosophila