Clear Sky Science · nl
In situ-architectuur van plasmodesmata in Physcomitrium patens opgelost met cryo-elektronentomografie
Piepkleine bruggen die plantencellen laten communiceren
Planten lijken stil, maar binnen hun weefsels wisselen cellen voortdurend signalen en voedingsstoffen uit. Dit verkeer moet door stijve celwanden heen, wat een vraag oproept: hoe blijven naburige cellen verbonden zonder grote openingen in hun wanden achter te laten? Deze studie zoomt in op de piepkleine kanalen die dit probleem oplossen in een mos en onthult hoe hun vorm en interne inrichting bepalen wanneer cellen contact houden en wanneer ze de deur sluiten.
Verborgen doorgangen in de plantenwand
Plantencellen zijn verbonden door microscopische tunnels die door hun gedeelde wanden lopen en directe bruggen vormen tussen het vocht- en membraansysteem van de ene cel en de volgende. In het mos Physcomitrium patens gebruikten de auteurs een cryogene afbeeldingsmethode die weefsel zo snel vriest dat water geen ijskristallen vormt. Vervolgens verzamelden ze driedimensionale beelden van deze bruggen in intact weefsel. De beelden tonen een eenvoudige maar opvallende opbouw: elk kanaal is bekleed door het buitenste celmembraan en bevat een dunnere binnenbuis die afkomstig is van het interne membraannetwerk van de cel. De nauwe ruimte tussen buitenwand en binnenbuis vormt een mouw waarlangs moleculen van cel naar cel kunnen bewegen, maar de breedte ervan varieert langs het kanaal en is het nauwst bij de openingen aan beide zijden.
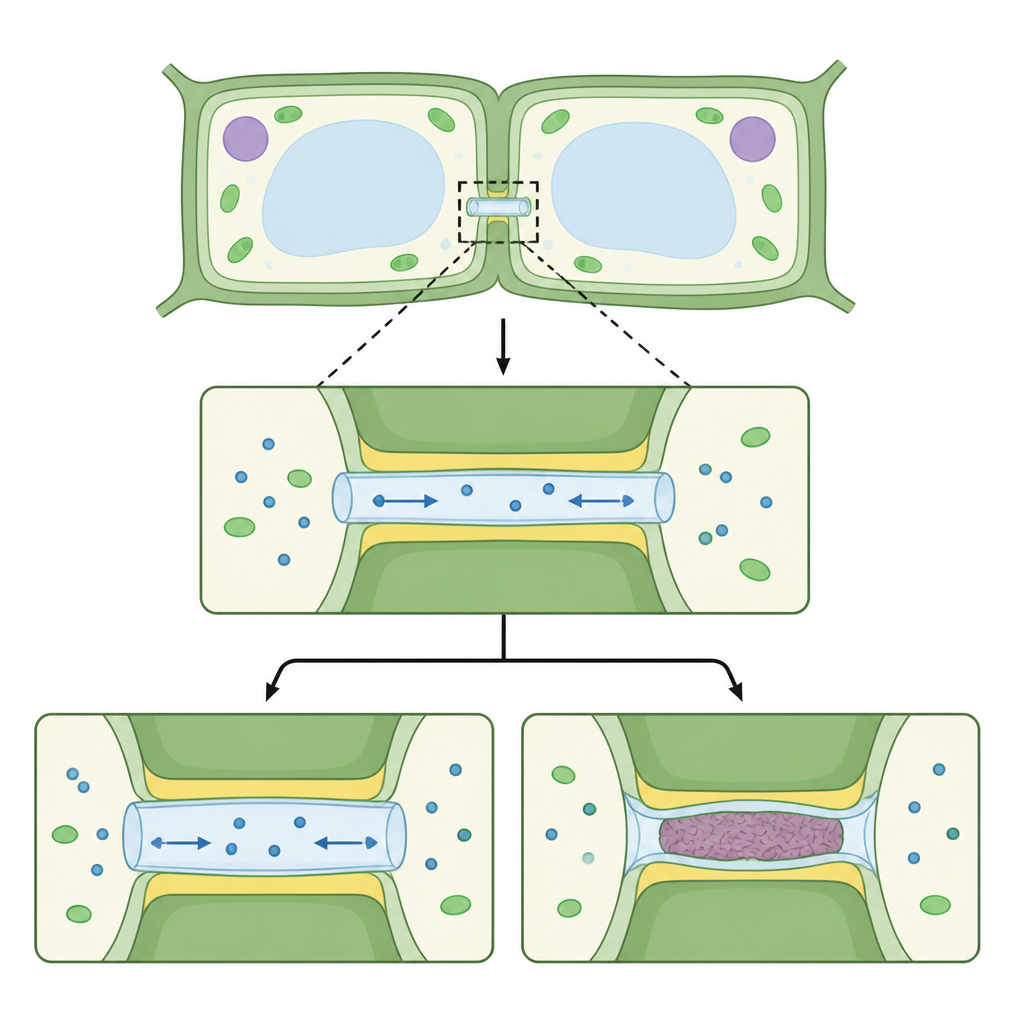
Hoe planten de passages verwijden of afsluiten
Planten passen aan hoe gemakkelijk moleculen deze bruggen passeren, en dit werk koppelt die regeling aan veranderingen in het omringende wandmateriaal. Het team onderzocht drie situaties in mosfilamenten: normaal weefsel, weefsel behandeld met het stresshormoon abscisinezuur en planten die zo zijn gemodificeerd dat ze een enzym overproduceren dat een wandpolymeer, callose, verwijdert. Wanneer abscisinezuur werd toegevoegd, vormden zich groffe, korrelige afzettingen rond de 'necks' van de kanalen. In veel gevallen sneden deze afzettingen de verbinding volledig af, zodat de binnenbuis, het buitenmembraan en de vloeistofmouw van beide cellen werden losgesneden en in de wand begraven achterbleven. Daarentegen werden de kanalen, wanneer callose actief werd verwijderd, korter en wijder over hun lengte. Deze veranderingen komen overeen met fysische modellen die voorspellen dat bredere, kortere tunnels moleculen gemakkelijker laten stromen, wat verklaart waarom deze gemodificeerde planten sterkere cel-tot-celuitwisseling tonen.
Een eiwitsteiger binnen het kanaal
Hoge-resolutieanalyse van de binnenbuis onthulde een verrassend intern skelet. Dicht bij de 'necks' van de kanalen is de buis omwikkeld door herhalende ringen van eiwit die eromheen draaien in een helisch rooster, vergelijkbaar met de windingen van een veer. Deze structuren verschijnen in zowel de belangrijkste mosweefsels als onder alle geteste omstandigheden, waarmee ze als kerncomponenten van het kanaalontwerp worden gemarkeerd. Door de gemeten vormen te vergelijken met computervoorspellingen van kandidaat-eiwitten die in deze bruggen zijn verrijkt, identificeerden de auteurs een familie genaamd Multiple C2 Domain and Transmembrane Proteins als de beste match. Hun modellen suggereren dat paren van deze eiwitten dimeriseren en tegen elkaar aan pakken om de helische mantel te vormen, met één uiteinde verankerd in het membraan van de buis en meerdere compacte domeinen die in elkaar grijpen om het geheel te stabiliseren.
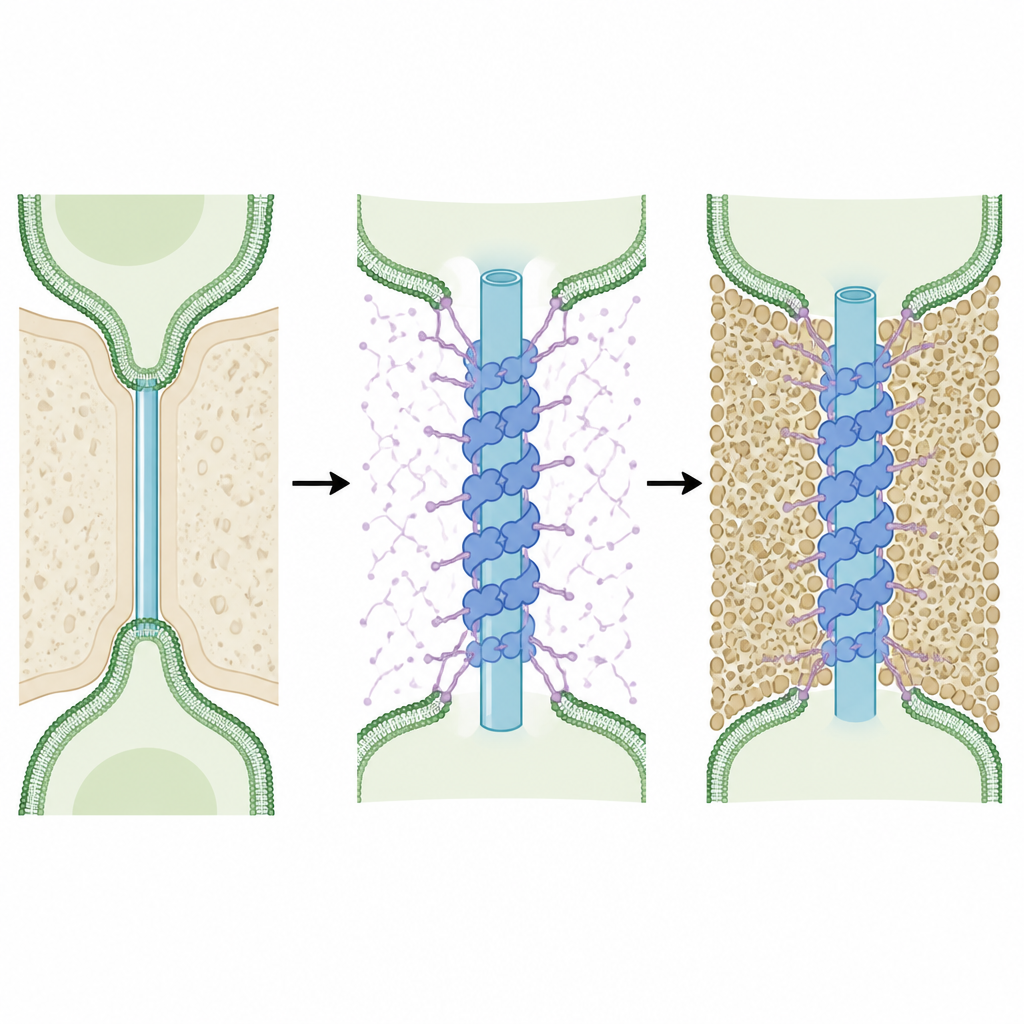
Flexibele strengen die het verkeer vormgeven
Elk eiwit in deze familie draagt ook een lang, slapper segment dat één van zijn domeinen met de rest van het molecuul verbindt. Voorspellingen en beeldanalyse wijzen uit dat deze flexibele verbindingsstukken naar buiten kunnen reiken vanaf de gecoate buis in de omliggende mouw en richting het buitenmembraan. De auteurs stellen voor dat veel van zulke linkers samen als ankers functioneren die de binnenbuis op zijn plaats houden, waardoor instorten of afbreken tijdens groei en afsluitgebeurtenissen wordt voorkomen. Omdat deze segmenten rijk zijn aan zowel positieve als negatieve ladingen en naar verwachting gedesordend blijven, kunnen ze de mouw ook vullen als een losse, dynamische mesh die beïnvloedt welke moleculen erdoorheen glippen — niet alleen op grootte maar ook op lading. Op deze manier kunnen het wandpolymeer callose, dat de mouwbreedte bepaalt, en de eiwitlinkers, die die ruimte vullen, samen werken om de connectiviteit tussen cellen fijn af te stemmen.
Waarom deze piepkleine bruggen ertoe doen
Deze studie levert een gedetailleerd beeld van hoe bruggen tussen plantencellen zijn opgebouwd en hoe ze reageren wanneer een mos stressgerelateerde toestanden ingaat. Ze toont dat een hormoonsignaal lokale herstructurering van de wand kan aansturen die kanalen volledig afsluit, terwijl een specifiek eiwitframe en zijn flexibele armen de structuur van de binnenbuis handhaven en helpen bij het bepalen van de regels voor moleculaire doorgang. Voor de niet-specialist is de kernboodschap dat plantencellen verbonden zijn door hooggeorganiseerde, aanpasbare deuropeningen waarvan de architectuur centraal staat voor hoe weefsels groeien, hulpbronnen delen en reageren op een veranderende omgeving.
Bronvermelding: Dickmanns, M., Pöge, M., Xu, P. et al. In situ architecture of plasmodesmata in Physcomitrium patens resolved by cryo-electron tomography. Nat. Plants 12, 1051–1061 (2026). https://doi.org/10.1038/s41477-026-02294-9
Trefwoorden: plasmodesmata, celcommunicatie bij planten, cryo-elektronentomografie, celwand, eiwitsteigers