Clear Sky Science · fr
Perte en plusieurs étapes des capacités catalytiques et de liaison des ligands de la purine nucléoside phosphorylase hexamérique
Pourquoi l’« usure » des enzymes compte
Les enzymes sont de petites machines moléculaires qui maintiennent chaque cellule en vie, des bactéries intestinales aux cellules immunitaires humaines. On a tendance à les voir comme soit fonctionnelles soit cassées, un peu comme une ampoule allumée ou éteinte. Cet article montre que, du moins pour une enzyme importante de la bactérie Escherichia coli, la réalité est beaucoup plus subtile. L’enzyme traverse progressivement une série d’états « à moitié fonctionnels » : elle peut encore se lier à ses partenaires moléculaires même après avoir largement perdu la capacité d’accélérer la réaction chimique. Comprendre ce cycle de vie caché modifie la façon dont les scientifiques mesurent les enzymes et pourrait aider à concevoir de meilleurs médicaments qui les ciblent.
Une machine clé de recyclage dans les cellules
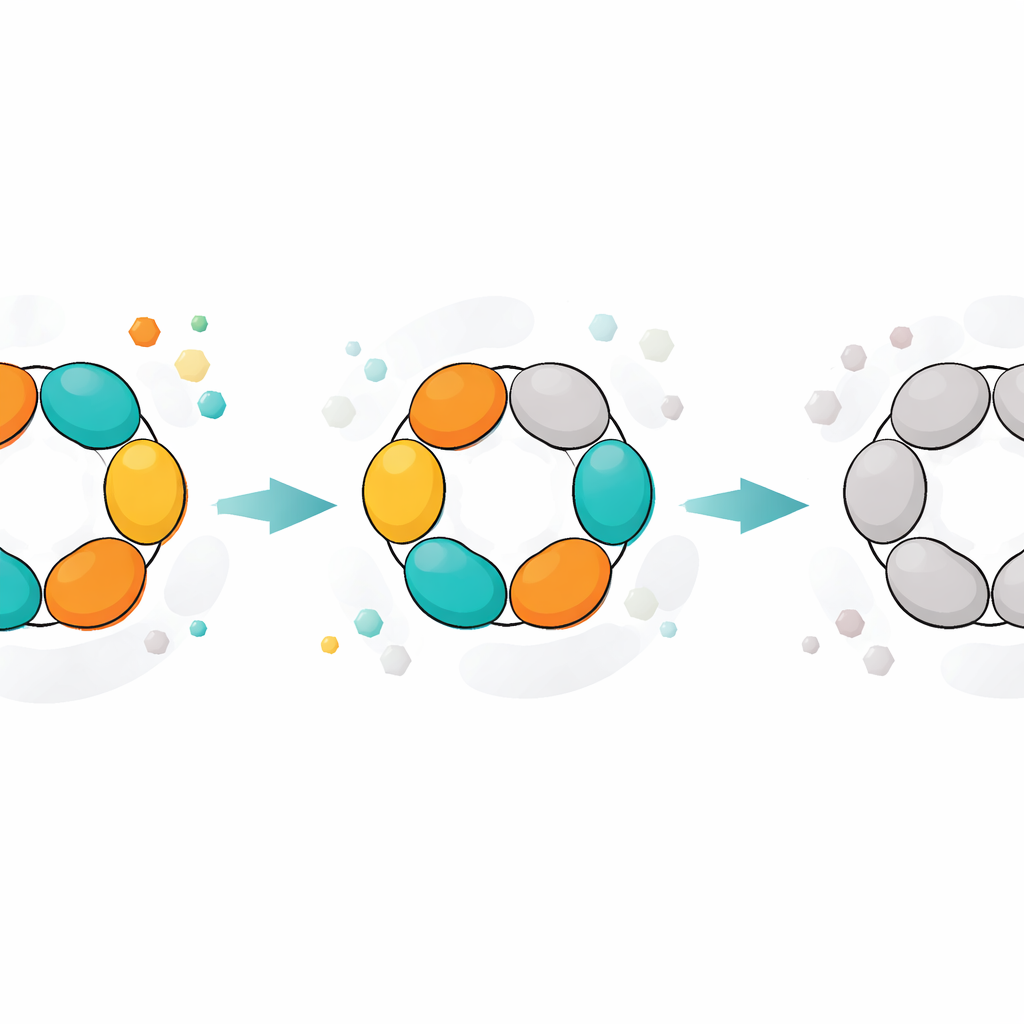
L’enzyme étudiée ici, la purine nucléoside phosphorylase (PNP), aide à recycler les blocs de construction de l’ADN et de l’ARN plutôt que de laisser les cellules les jeter inutilement. Les bactéries comme E. coli dépendent fortement de cette voie de recyclage, et certains agents pathogènes en dépendent presque complètement. La PNP a aussi une importance médicale chez l’humain : bloquer la version humaine peut atténuer certains lymphocytes et est déjà utilisé pour traiter des cancers du système immunitaire. Pour ces raisons, la PNP et ses inhibiteurs ont été étudiés comme cibles thérapeutiques contre le cancer, les maladies auto‑immunes et les infections. L’enzyme d’E. coli présente une architecture frappante : six unités protéiques identiques forment un hexamère en anneau, organisé en trois paires (dimères). Chaque dimère héberge les poches où se déroule la chimie, et ces poches doivent s’ouvrir et se fermer de façon coordonnée pour que la réaction ait lieu.
Une énigme : pur, bien comporté et pourtant perdant de la puissance
Au fil d’années d’expériences, les auteurs ont remarqué quelque chose d’étrange. Des échantillons soigneusement purifiés de PNP d’E. coli, qui paraissaient parfaitement propres sur des gels protéiques standards, ne se comportaient pas comme des copies identiques de la même machine. Leur « activité spécifique » – la vitesse à laquelle ils traitaient un substrat naturel standard, l’inosine – pouvait commencer très élevée puis chuter en plusieurs phases sur des jours ou des mois, même lorsque la protéine était conservée congelée. Différentes conditions de stockage et tampons modifiaient la vitesse et le motif de ce déclin. Plus surprenant encore, certains échantillons ayant presque complètement perdu leur activité vis‑à‑vis de l’inosine pouvaient encore traiter efficacement un substrat modifié, la 7‑méthylguanosine, qui imite étroitement l’état de transition fugitif de la réaction. Autrement dit, l’enzyme pouvait toujours retenir ses partenaires mais était devenue très mauvaise pour accomplir la partie difficile du travail.
Lier sans accomplir la réaction
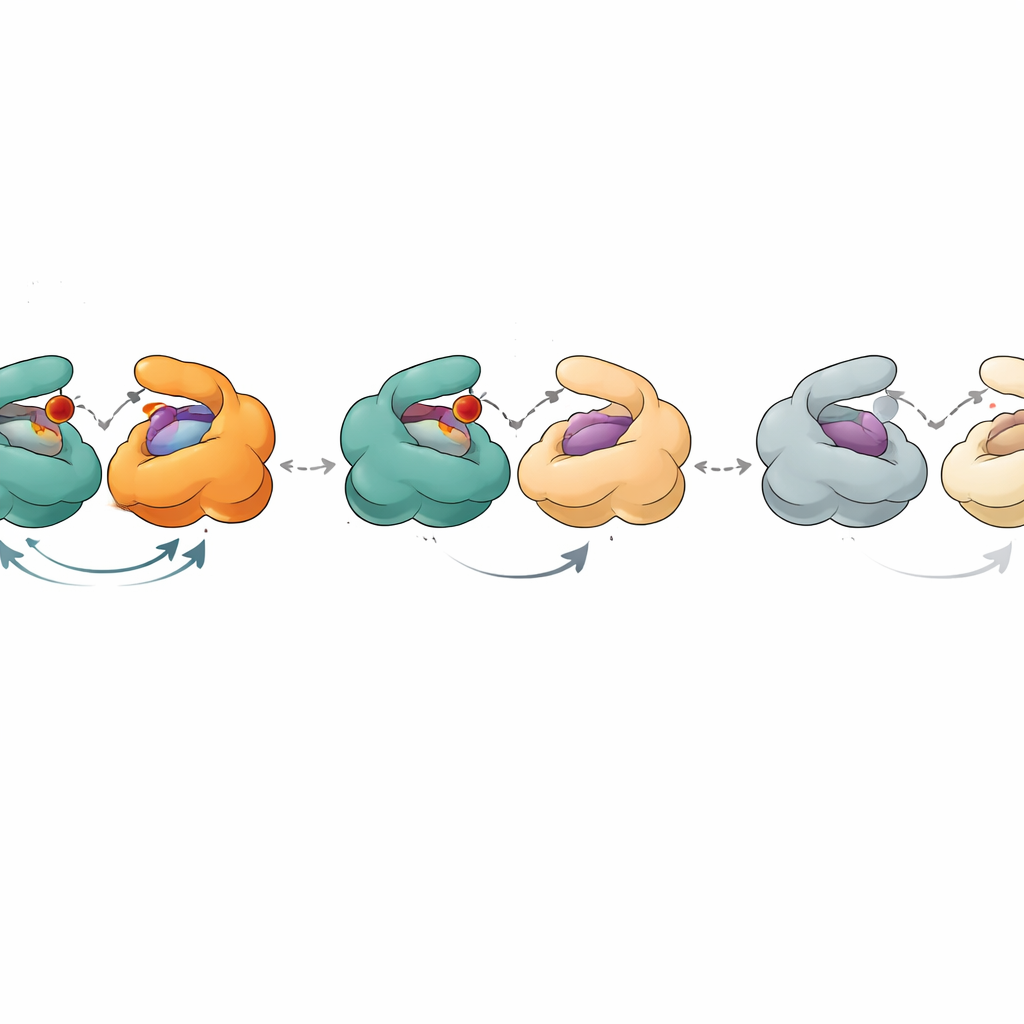
Pour approfondir cela, les chercheurs ont mesuré la capacité des échantillons de PNP vieillissants à se lier au phosphate (un co‑substrat) et à divers molécules de type nucléoside en utilisant des techniques sensibles comme la calorimétrie et la titration de fluorescence. La pratique standard suppose que toute protéine inactive dans un échantillon « pur » ne lie pas non plus les ligands, elle est donc ignorée dans l’analyse. Ici, cette hypothèse s’est effondrée. En combinant des mesures d’activité et des courbes de liaison, les auteurs ont pu estimer quelle fraction de chaque échantillon était catalytiquement active et quelle fraction se liait encore aux ligands. Ils ont constaté que la fraction compétente pour la liaison était systématiquement plus grande que la fraction compétente pour la catalyse. De plus, les constantes de dissociation apparentes – nombres décrivant la force de liaison des ligands – évoluaient de manière systématique à mesure que l’enzyme vieillissait. Les données ne pouvaient s’expliquer que si les six sites actifs de l’hexamère n’étaient pas tous équivalents et si des états intermédiaires, toujours capables de lier mais peu performants en catalyse, s’accumulaient avec le temps.
Instantanés structuraux d’une machine vieillissante
Les structures cristallographiques de la PNP ont fourni une image physique de ces états. Dans la forme la plus active, les trois dimères présentent chacun un site actif ouvert et un site actif fermé, et les six sous‑unités coopèrent dans un cycle synchronisé d’ouverture et de fermeture en « bascule ». En vieillissant, un dimère cesse effectivement de participer : ses deux sites restent ouverts et ne se ferment plus suffisamment pour soutenir la réaction normale, bien qu’ils puissent encore retenir des ligands. Les deux dimères restants fonctionnent toujours, mais moins efficacement. Dans des états encore plus avancés, aucun des dimères ne peut se fermer correctement. Dans ces formes ouvertes, des substrats naturels comme l’inosine ne sont plus transformés, mais des substrats imitant l’état de transition qui portent déjà une charge positive clé peuvent encore être dégradés, contournant le besoin de mouvements structuraux précis. Une enzyme apparentée de Helicobacter pylori, qui a une forme globale très similaire, a montré un déclin plus progressif et moins par étapes, ce qui suggère une coopération plus faible entre ses sous‑unités et explique sa activité maximale plus faible.
Ce que cela signifie pour la biologie et la conception de médicaments
Ce travail révèle que, pour la PNP hexamérique d’E. coli, l’inactivation n’est pas un simple événement marche‑arrêt. L’enzyme progresse à travers des états intermédiaires bien définis dans lesquels certains dimères sont pleinement actifs, certains ne peuvent que lier des ligands et d’autres sont essentiellement inactifs. De ce fait, la force de liaison mesurée pour des candidats‑médicaments ou des substrats dépend de l’« âge » de la préparation enzymatique. Pour les chercheurs, cela signifie que l’activité spécifique n’est pas un détail annexe mais une partie essentielle de l’interprétation des données de liaison et d’inhibition. Plus généralement, l’étude montre comment des enzymes multi‑sous‑unités peuvent perdre leur coordination progressivement, comme une machine complexe qui tourne encore mais ne délivre plus sa pleine puissance. Reconnaître et caractériser ces états intermédiaires pourrait affiner notre compréhension du fonctionnement enzymatique dans les cellules et améliorer la fiabilité des expériences qui guident le développement de nouvelles thérapies.
Citation: Narczyk, M., Bzowska, A. Multistep loss of catalytic and ligand binding abilities of hexameric purine nucleoside phosphorylase. Sci Rep 16, 11553 (2026). https://doi.org/10.1038/s41598-026-41204-z
Mots-clés: vieillissement des enzymes, purine nucléoside phosphorylase, enzymes allostériques, liaison enzyme–ligand, coopérativité des protéines