Clear Sky Science · es
Pérdida en varios pasos de las capacidades catalíticas y de unión a ligandos de la purina nucleósido fosforilasa hexamérica
Por qué importa el “desgaste” enzimático
Las enzimas son pequeñas máquinas moleculares que mantienen vivas a todas las células, desde las bacterias intestinales hasta las células inmunitarias humanas. Tendemos a pensar en ellas como si funcionaran o estuvieran rotas, como una bombilla que está encendida o apagada. Este artículo muestra que, al menos para una enzima importante de la bacteria Escherichia coli, la realidad es mucho más sutil. La enzima pasa gradualmente por una serie de estados “medio‑funcionales”: puede seguir agarrando a sus moléculas compañeras incluso después de haber prácticamente olvidado cómo acelerar la reacción química. Entender este ciclo de vida oculto cambia la forma en que los científicos miden las enzimas y podría ayudar a diseñar mejores fármacos que las tengan como diana.
Una máquina clave de reciclaje en las células
La enzima estudiada aquí, la purina nucleósido fosforilasa (PNP), ayuda a reciclar los bloques de construcción del ADN y ARN en lugar de que las células los desperdicien. Bacterias como E. coli dependen en gran medida de esta vía de reciclaje, y algunos microbios patógenos dependen de ella casi por completo. La PNP también es médicamente importante en humanos: bloquear la versión humana puede reducir ciertas células inmunitarias y ya se utiliza en el tratamiento de cánceres del sistema inmune. Por ello, la PNP y sus inhibidores se han explorado como dianas terapéuticas para cáncer, enfermedades autoinmunes e infecciones. La enzima de E. coli tiene una arquitectura llamativa: seis unidades proteicas idénticas forman un hexámero en forma de anillo, organizado como tres pares (dímeros). Cada dímero alberga los bolsillos donde ocurre la química, y esos bolsillos deben abrirse y cerrarse de forma coordinada para que la reacción avance.
Un enigma: puro, bien comportado y aun así perdiendo potencia
A lo largo de muchos años de experimentos, los autores notaron algo extraño. Muestras de PNP de E. coli cuidadosamente purificadas, que parecían perfectamente limpias en geles proteicos estándar, no se comportaban como copias idénticas de la misma máquina. Su “actividad específica”, es decir, la velocidad con la que procesaban un sustrato natural estándar llamado inosina, podía comenzar muy alta y luego caer en varias fases durante días o meses, incluso cuando la proteína se mantenía congelada. Diferentes condiciones de almacenamiento y tampones cambiaban la velocidad y el patrón de este descenso. Aún más sorprendente, algunas muestras que casi habían perdido por completo la actividad frente a la inosina podían seguir procesando eficientemente un sustrato modificado, la 7‑metilguanosina, que imita de cerca el efímero estado de transición de la reacción. En otras palabras, la enzima todavía podía sostener a sus socios pero se había vuelto muy mala en la parte difícil del trabajo.
Unir sin hacer el trabajo
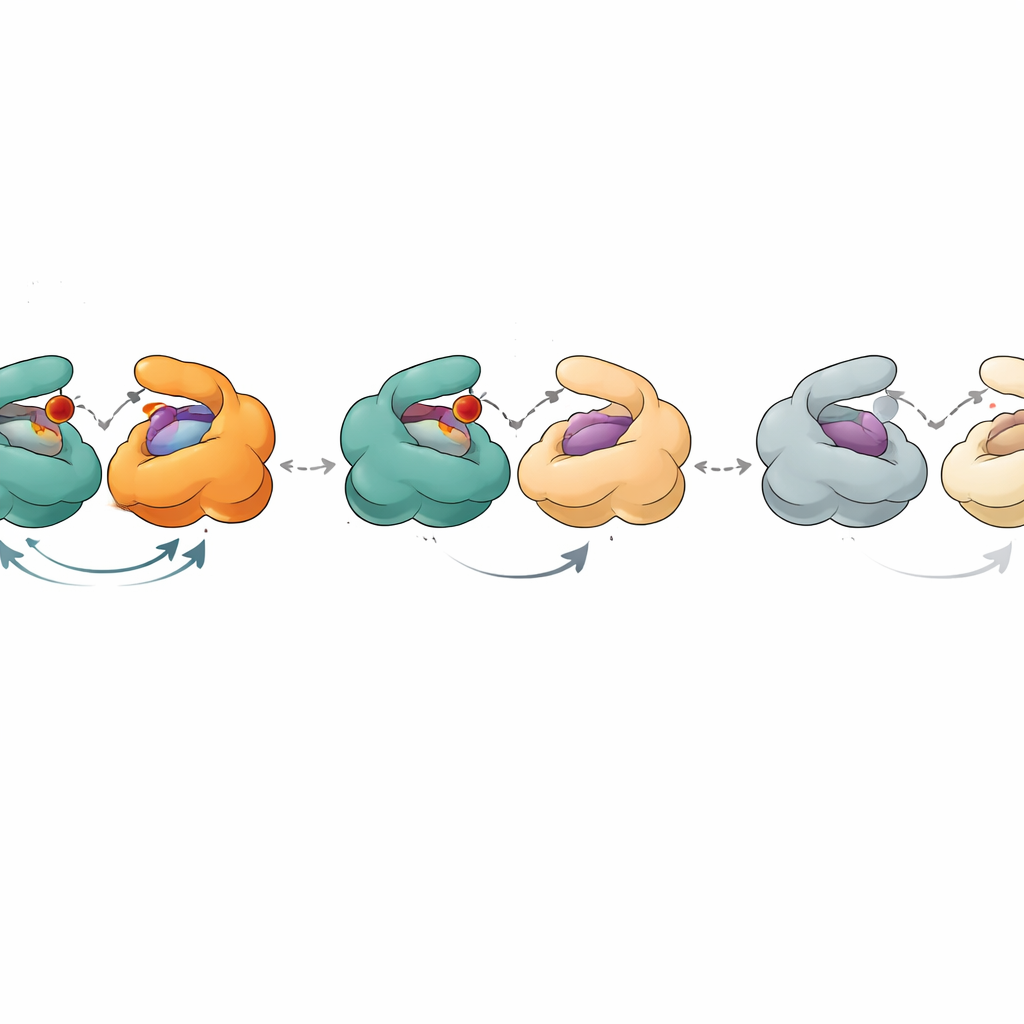
Para profundizar en esto, los investigadores midieron cuánto unían las muestras envejecidas de PNP al fosfato (un cosustrato) y a varias moléculas similares a nucleósidos usando técnicas sensibles como calorimetría y titulaciones de fluorescencia. La práctica habitual asume que cualquier proteína inactiva en una muestra “pura” tampoco se une a ligandos, por lo que se la ignora en el análisis. Aquí, esa suposición falló. Combinando medidas de actividad con curvas de unión, los autores pudieron estimar qué fracción de cada muestra era catalíticamente activa y qué fracción seguía uniendo ligandos. Encontraron que la fracción competente para la unión era sistemáticamente mayor que la fracción competente catalíticamente. Además, las constantes aparentes de disociación —números que describen cuán fuertemente se unen los ligandos— cambiaron de forma sistemática a medida que la enzima envejecía. Los datos solo podían explicarse si los seis sitios activos en el hexámero no eran todos equivalentes y si se acumulaban estados intermedios, capaces aún de unir pero pobres en catálisis, con el tiempo.
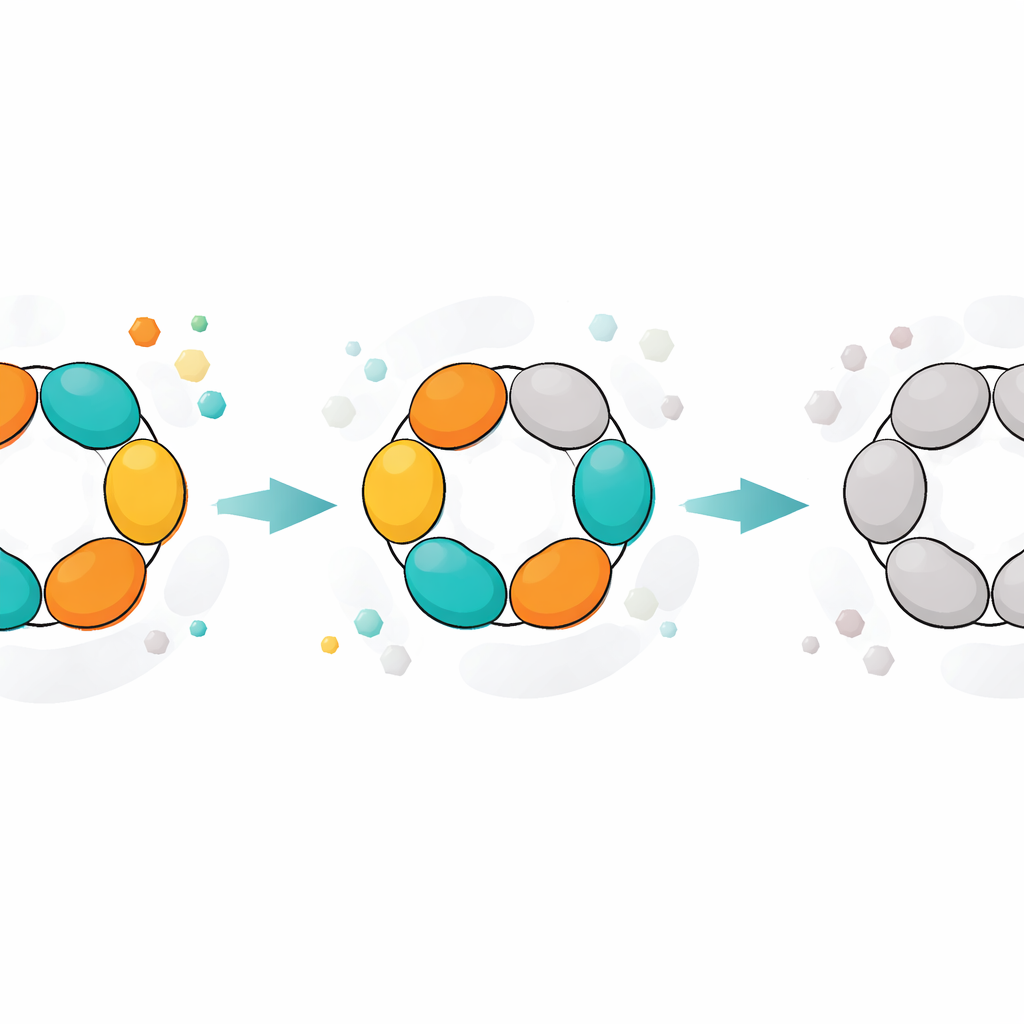
Instantáneas estructurales de una máquina que envejece
Las estructuras cristalográficas de la PNP proporcionaron una imagen física de estos estados. En la forma más activa, los tres dímeros muestran un sitio activo abierto y otro cerrado, y las seis subunidades cooperan en un ciclo sincronizado de «flip‑flop» de apertura y cierre. A medida que la enzima envejece, un dímero deja de participar efectivamente: ambos de sus sitios permanecen abiertos y ya no se cierran con la suficiente intensidad para soportar la reacción normal, aunque todavía pueden retener ligandos. Los dos dímeros restantes siguen funcionando, pero con menor eficiencia. En estados aún más tardíos, ninguno de los dímeros puede cerrarse correctamente. En estas formas abiertas, sustratos naturales como la inosina ya no se procesan, pero sustratos que imitan el estado de transición y que ya portan una carga positiva clave pueden seguir degradándose, evitando la necesidad de movimientos estructurales precisos. Una enzima relacionada de Helicobacter pylori, que tiene una forma global muy similar, mostró un declive más suave y menos escalonado, lo que sugiere una cooperatividad más débil entre sus subunidades y explica su menor actividad máxima.
Qué significa esto para la biología y el diseño de fármacos
Este trabajo revela que, para la PNP hexamérica de E. coli, la inactivación no es un simple evento de encendido‑apagado. La enzima progresa a través de estados intermedios bien definidos en los que algunos dímeros son completamente activos, otros solo son capaces de unir ligandos y otros están esencialmente inactivos. Debido a esto, la fuerza de unión medida de candidatos a fármacos o de sustratos depende de cuánto esté “envejecida” la preparación enzimática. Para los investigadores, esto significa que la actividad específica no es un mero comentario accesorio sino una parte esencial para interpretar datos de unión e inhibición. Más ampliamente, el estudio muestra cómo enzimas multimericas pueden perder coordinación de forma gradual, como una máquina compleja que sigue girando pero ya no entrega su rendimiento completo. Reconocer y caracterizar estos estados intermedios podría afinar nuestra comprensión de la función enzimática en las células y mejorar la fiabilidad de los experimentos que guían nuevas terapias.
Cita: Narczyk, M., Bzowska, A. Multistep loss of catalytic and ligand binding abilities of hexameric purine nucleoside phosphorylase. Sci Rep 16, 11553 (2026). https://doi.org/10.1038/s41598-026-41204-z
Palabras clave: envejecimiento enzimático, purina nucleósido fosforilasa, enzimas alostéricas, unión enzima–ligando, cooperatividad proteica