Clear Sky Science · en
Multistep loss of catalytic and ligand binding abilities of hexameric purine nucleoside phosphorylase
Why enzyme “wear and tear” matters
Enzymes are tiny molecular machines that keep every cell alive, from gut bacteria to human immune cells. We tend to think of them as either working or broken, like a lightbulb that is on or off. This paper shows that, at least for one important enzyme from the bacterium Escherichia coli, reality is far subtler. The enzyme gradually passes through a series of “half‑working” states: it can still grab onto its partner molecules even after it has largely forgotten how to speed up the chemical reaction. Understanding this hidden life cycle changes how scientists measure enzymes and could help in designing better drugs that target them.
A key recycling machine in cells
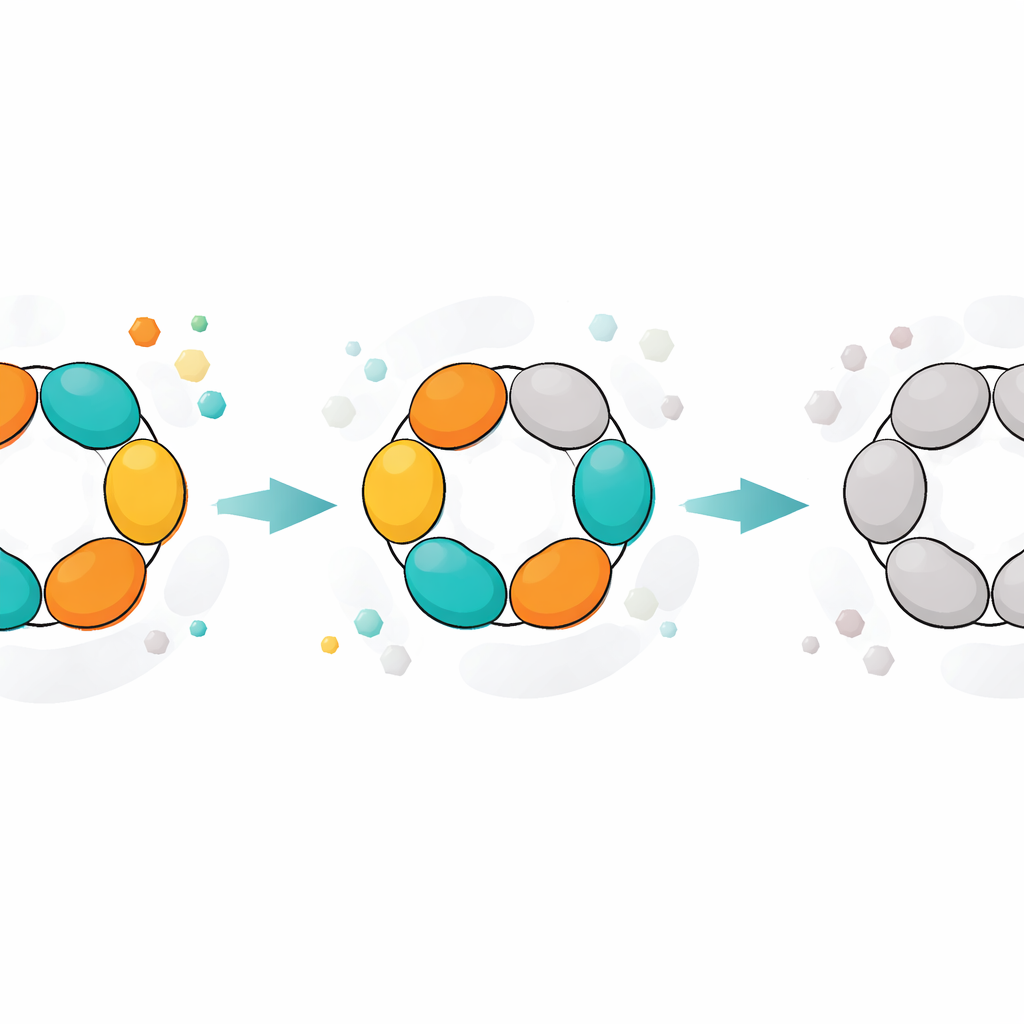
The enzyme studied here, purine nucleoside phosphorylase (PNP), helps recycle the building blocks of DNA and RNA rather than letting cells wastefully throw them away. Bacteria such as E. coli rely heavily on this recycling pathway, and some disease‑causing microbes depend on it almost completely. PNP is also medically important in humans: blocking the human version can damp down certain immune cells and is already used to treat cancers of the immune system. Because of this, PNP and its inhibitors have been explored as drug targets for cancer, autoimmune disease, and infections. The E. coli enzyme has a striking architecture: six identical protein units form a ring‑like hexamer, organized as three pairs (dimers). Each dimer hosts the pockets where chemistry happens, and these pockets must open and close in a coordinated way for the reaction to proceed.
A puzzle: pure, well‑behaved and yet losing power
Over many years of experiments the authors noticed something odd. Carefully purified samples of E. coli PNP, which looked perfectly clean on standard protein gels, did not behave as identical copies of the same machine. Their “specific activity” – how fast they processed a standard natural substrate called inosine – could start very high and then drop in multiple phases over days or months, even when the protein was kept frozen. Different storage conditions and buffers changed the rate and pattern of this decline. Even more surprising, some samples that had almost completely lost activity toward inosine could still efficiently process a modified substrate, 7‑methylguanosine, which closely mimics the fleeting transition state of the reaction. In other words, the enzyme could still hold its partners but had become very poor at doing the hard part of the job.
Binding without doing the job
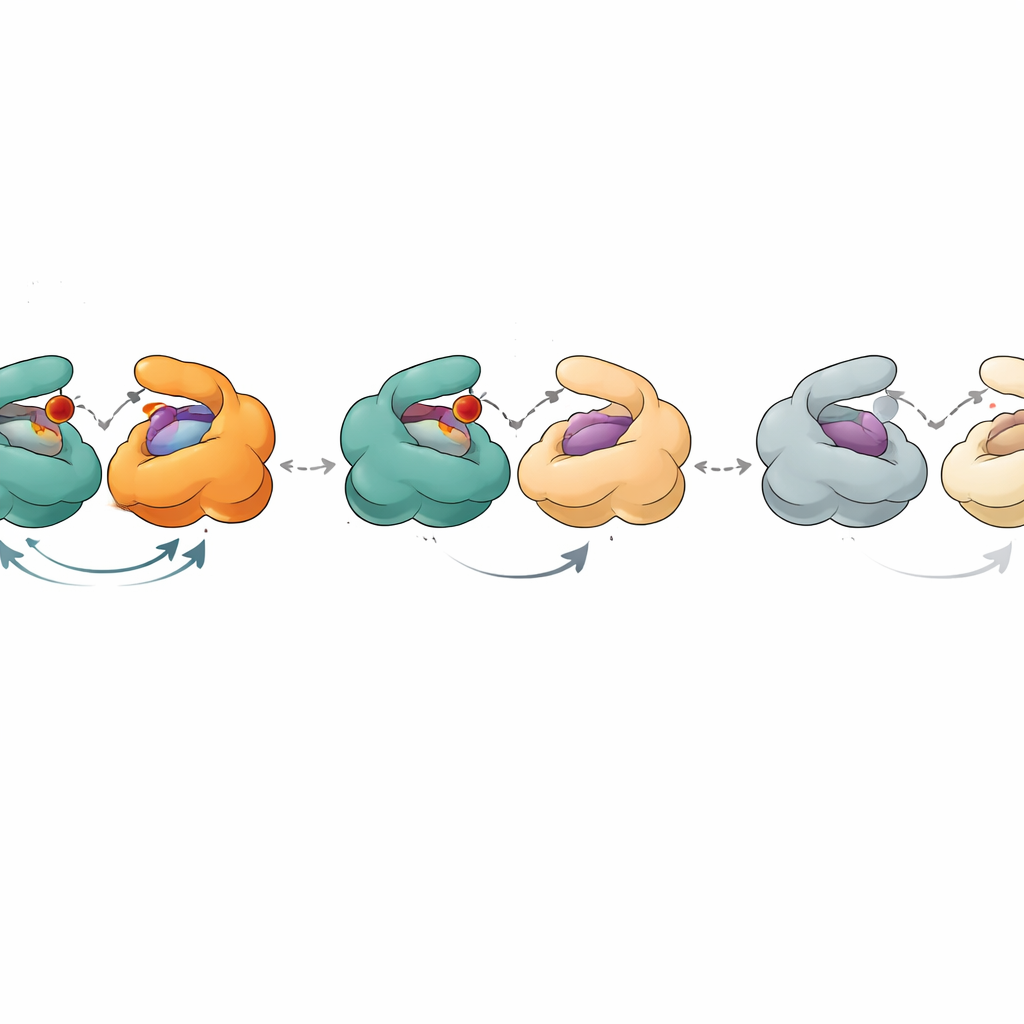
To probe this more deeply, the researchers measured how well aging PNP samples bound to phosphate (a co‑substrate) and to various nucleoside‑like molecules using sensitive techniques such as calorimetry and fluorescence titration. Standard practice assumes that any inactive protein in a “pure” sample also fails to bind ligands, so it is ignored in the analysis. Here, that assumption broke down. By combining activity measurements with binding curves, the authors could estimate how much of each sample was catalytically active and how much still bound ligands. They found that the binding‑competent fraction was consistently larger than the catalytically competent fraction. Moreover, the apparent dissociation constants – numbers that describe how tightly ligands bind – shifted in a systematic way as the enzyme aged. The data could only be explained if the six active sites in the hexamer were not all equivalent and if intermediate states, still able to bind but poor at catalysis, accumulated over time.
Structural snapshots of an aging machine
Crystal structures of PNP provided a physical picture of these states. In the most active form, all three dimers show one open and one closed active site, and the six subunits cooperate in a synchronized “flip‑flop” cycle of opening and closing. As the enzyme ages, one dimer effectively drops out: both of its sites remain open and no longer close tightly enough to support the normal reaction, even though they can still hold ligands. The remaining two dimers still function, but less efficiently. In even later states, none of the dimers can close properly. In these open forms, natural substrates like inosine are no longer processed, but transition‑state‑like substrates that already carry a key positive charge can still be broken down, bypassing the need for precise structural movements. A related enzyme from Helicobacter pylori, which has a very similar overall shape, showed a smoother, less step‑wise decline, suggesting weaker cooperation between its subunits and explaining its lower maximal activity.
What this means for biology and drug design
This work reveals that for hexameric PNP from E. coli, inactivation is not a simple on‑off event. The enzyme progresses through well‑defined intermediate states in which some dimers are fully active, some are only able to bind ligands, and others are essentially idle. Because of this, the measured binding strength of drug candidates or substrates depends on how “aged” the enzyme preparation is. For researchers, this means that specific activity is not just a side note but an essential part of interpreting binding and inhibition data. More broadly, the study shows how multi‑subunit enzymes can lose coordination gradually, much like a complex machine that keeps turning but no longer delivers its full output. Recognizing and characterizing these in‑between states could sharpen our understanding of enzyme function in cells and improve the reliability of experiments that guide new therapies.
Citation: Narczyk, M., Bzowska, A. Multistep loss of catalytic and ligand binding abilities of hexameric purine nucleoside phosphorylase. Sci Rep 16, 11553 (2026). https://doi.org/10.1038/s41598-026-41204-z
Keywords: enzyme aging, purine nucleoside phosphorylase, allosteric enzymes, enzyme–ligand binding, protein cooperativity