Clear Sky Science · fr
Les anneaux MCM archéens et eucaryotes ouvrent séquentiellement l’ADN pour initier la réplication
Comment nos cellules commencent à copier l’ADN
Chaque fois qu’une cellule se divise, elle doit copier l’ensemble de son livre d’instructions génétiques avec une précision extraordinaire. Ce processus de copie commence par un premier geste délicat : une petite portion de la double hélice d’ADN doit s’ouvrir pour que la machinerie de copie puisse accéder aux brins. Cette étude révèle, au niveau moléculaire, comment une machinerie protéique clé en forme d’anneau, présente chez les organismes simples comme complexes, réalise ce premier acte d’ouverture, préparant ainsi le terrain pour une réplication fidèle de l’ADN.

Le moteur de copie de l’ADN dans tous les domaines du vivant
La réplication de l’ADN repose sur des enzymes appelées hélicases qui séparent les deux brins de la double hélice, créant des matrices pour la synthèse de l’ADN nouveau. Chez les bactéries, un complexe protéique ouvre d’abord l’ADN, puis une hélicase annulaire distincte est chargée. En revanche, chez les archées et les eucaryotes, une hélicase connue sous le nom de complexe MCM est chargée sur de l’ADN double brin intact puis n’est activée que plus tard. Cet assemblage annulaire, composé de six sous‑unités protéiques apparentées, doit d’une manière ou d’une autre transformer une hélice entièrement appariée en une structure partiellement ouverte que d’autres enzymes pourront étendre en une fourche de réplication complète.
Instantanés de l’ADN au tout début de son ouverture
Les chercheurs ont utilisé la cryo‑microscopie électronique à haute résolution pour capturer de nombreux instantanés d’un anneau MCM d’archée entourant un court segment d’ADN. Ils ont observé deux arrangements principaux. Dans l’un, l’anneau forme deux étages alignés et entoure lâchement l’ADN parfaitement apparié, n’établissant que des contacts légers. Dans l’autre, les étages sont déplacés l’un par rapport à l’autre et l’étage inférieur saisit un des brins d’ADN beaucoup plus fermement. Dans cette configuration décalée, une partie de l’ADN près d’une extrémité n’est plus appariée et a fondu en simples brins, alors que l’ADN de départ était entièrement double brin.
Une petite cale aromatique qui écarte l’ADN
Un examen plus attentif a montré que trois sous‑unités voisines dans l’anneau actif utilisent de petites boucles projetées pour contacter l’un des brins d’ADN. Chaque boucle porte un groupe chimique plat appelé anneau aromatique qui s’empile contre le sucre et la base de l’ADN comme une cale. Quand un ou deux des interstices entre sous‑unités se resserrent, ces cales s’insèrent dans le sillon mineur de l’ADN et décollent deux paires de bases. Quand un troisième interstice se resserre, quatre paires de bases sont fondues. Ces étapes de resserrement sont liées à la liaison de molécules d’ATP en sites spécifiques entre sous‑unités, ce qui suggère une séquence où la liaison de l’ATP entraîne des augmentations discontinues de l’ouverture locale de l’ADN.
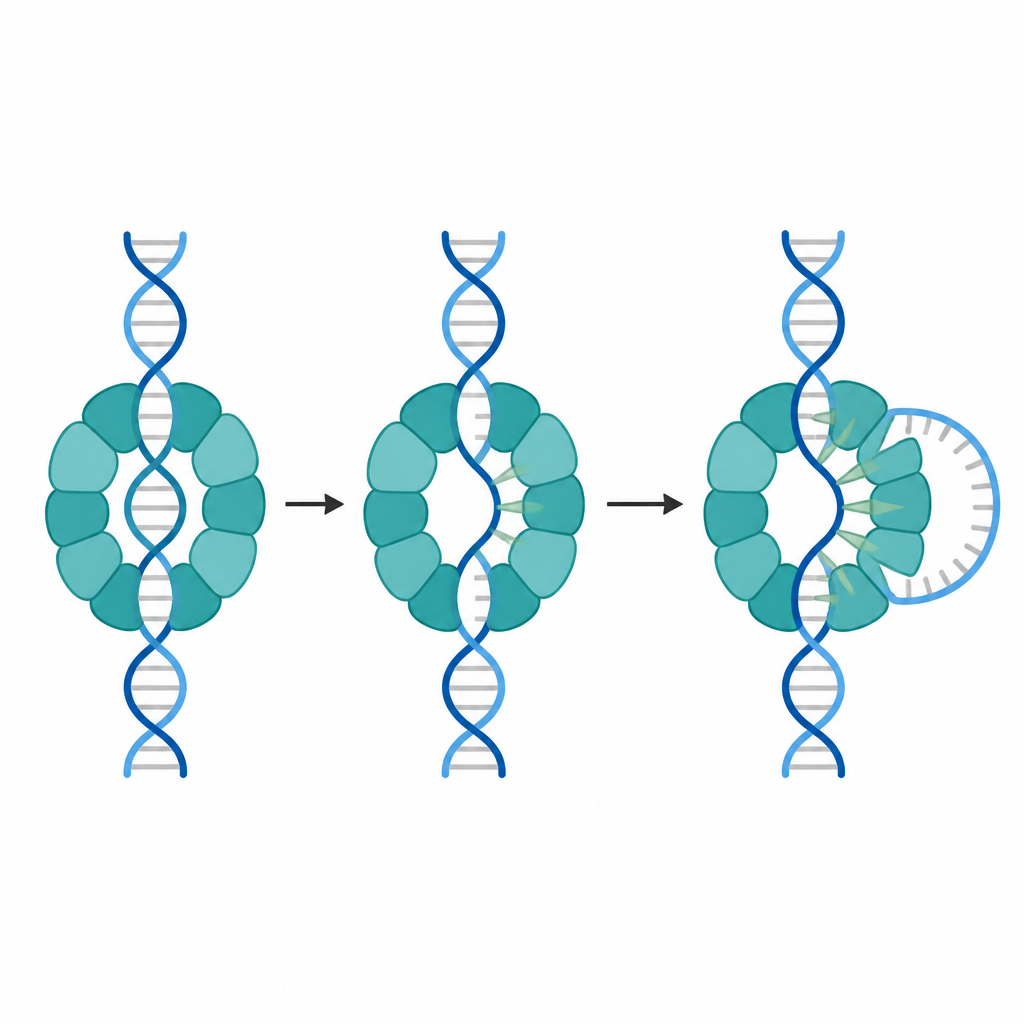
Un mouvement d’ouverture universel partagé entre espèces et virus
Pour tester si ce mécanisme est spécifique au système archéen ou s’il s’applique de façon générale, l’équipe a comparé leurs structures à des dizaines de structures d’hélicases déjà résolues chez la levure, l’homme et des virus à ADN oncogènes. Ils ont constaté que les anneaux MCM eucaryotes adoptent également deux conformations globales stables : l’une qui maintient l’ADN entièrement apparié et l’autre qui place trois cales aromatiques équivalentes en position de fusion. Les hélicases virales du papillomavirus et du SV40 utilisent des groupes aromatiques étroitement apparentés en positions similaires pour ouvrir l’ADN de l’origine. Cette conservation suggère qu’un mécanisme de fusion basé sur des cales aromatiques est partagé par les archées, les eucaryotes et plusieurs virus à ADN.
Des premières paires de bases fondues aux fourches de réplication complètes
Ce travail étaye l’idée selon laquelle la liaison de l’ATP convertit un anneau MCM relâché en une forme active qui écarte seulement quelques paires de bases d’ADN à l’aide de ses cales aromatiques. D’autres facteurs cellulaires peuvent ensuite faire passer l’ADN le long de cette cale fixe, étendant la région fondue jusqu’à ce que les deux brins soient entièrement séparés et que l’hélicase n’entoure qu’un seul d’entre eux. En termes simples, l’étude explique comment un anneau moléculaire craque délicatement la fermeture éclair de l’ADN au moment et à l’endroit exacts, lançant le processus complexe de duplication du génome.
Citation: Rasouli, S., Myasnikov, A. & Enemark, E.J. Archaeal and eukaryotic MCM rings sequentially melt DNA for replication initiation. Nat Commun 17, 4681 (2026). https://doi.org/10.1038/s41467-026-70961-8
Mots-clés: initiation de la réplication de l’ADN, hélicase MCM, cale aromatique, cryo‑microscopie électronique, fusion de l’origine