Pourquoi notre cerveau se préoccupe de minuscules déplacements
Chaque fois que vous attrapez une balle, remarquez une voiture qui passe ou observez simplement des feuilles qui bougent dans le vent, votre système visuel résout discrètement un problème difficile : à partir d’un fouillis de motifs lumineux changeants sur la rétine, il doit déterminer dans quelle direction les objets se déplacent. Pendant des décennies, les scientifiques se sont appuyés sur quelques théories classiques pour expliquer comment les neurones détectent le mouvement. Cette étude utilise un apprentissage automatique inspiré biologiquement pour poser une question plus audacieuse : existe-t-il bien plus de façons pour de simples circuits neuronaux de percevoir le mouvement que nous ne l’imaginions jusqu’ici ?
Idées classiques sur la vision du mouvement
Les premiers travaux sur la vision du mouvement ont proposé que des neurones sensibles à la direction fonctionnent comme de petits calculateurs temporels. Dans un schéma traditionnel, les entrées provenant de points voisins dans l’espace sont comparées après qu’une d’entre elles a été retardée, de sorte que les signaux se déplaçant dans la direction préférée arrivent ensemble et s’additionnent, alors que le mouvement dans la direction opposée ne le fait pas. Un autre modèle classique oppose une excitation rapide à une inhibition retardée provenant du côté opposé du champ récepteur d’une cellule, annulant les réponses au mouvement dans la direction défavorisée. Ces mécanismes, bien qu’influents, se concentrent principalement sur les différences de timing et laissent ouverte la question de savoir si d’autres astuces de circuit pourraient atteindre le même résultat.
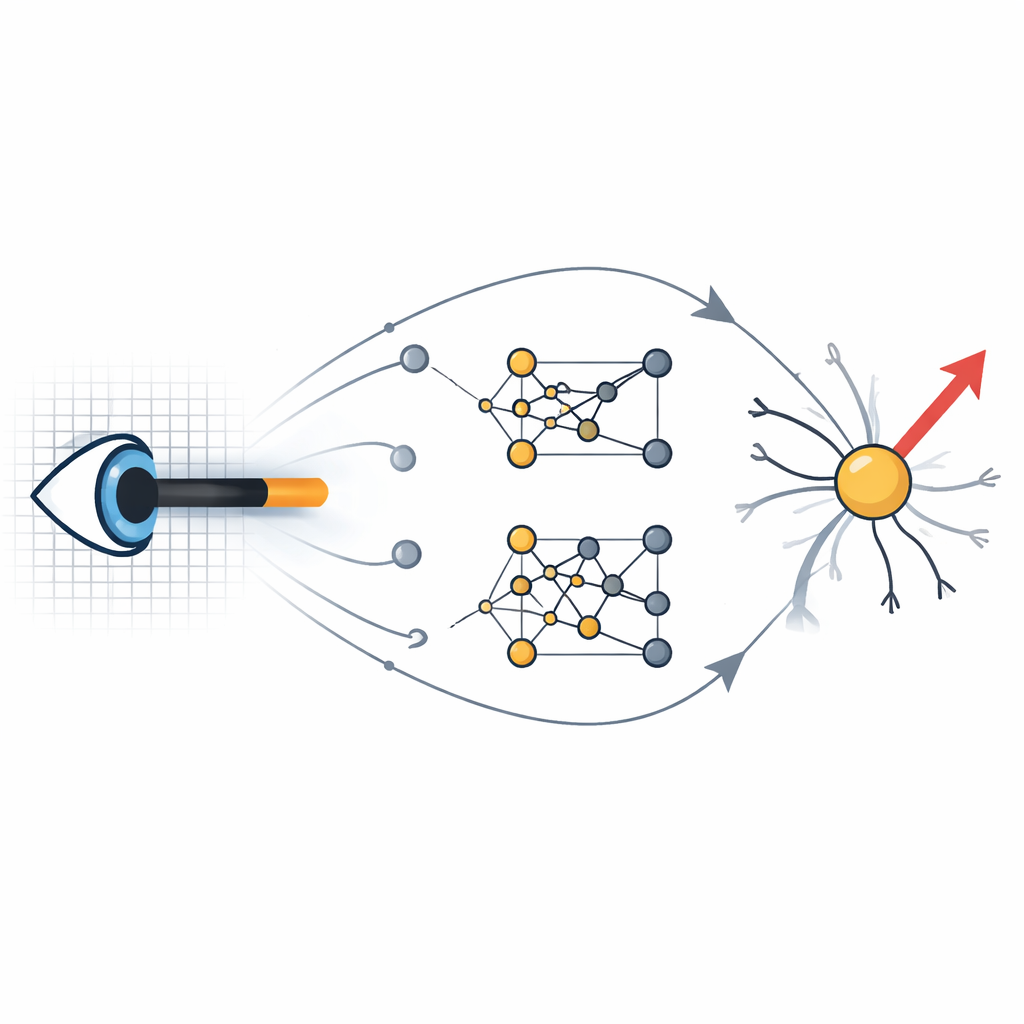
Laisser l’évolution explorer l’espace des circuits Figure 1.
Pour explorer cette question, l’auteur a construit des modèles informatiques détaillés de neurones de la rétine et du cortex visuel, puis a mis en place une recherche évolutionnaire. À partir de câblages aléatoires et de propriétés synaptiques aléatoires, un algorithme génétique a muté à plusieurs reprises les paramètres de circuit et sélectionné les modèles qui signalaient le mieux la bonne direction de barres en mouvement et de réseaux de grains défilants. Il est important de noter que les cellules d’entrée individuelles alimentant les détecteurs de mouvement n’étaient pas elles-mêmes sensibles à la direction ; seule leur organisation collective et leur dynamique pouvaient donner lieu à des sorties accordées. Cette exploration automatisée a mis au jour de nombreuses architectures de circuits différentes rivalisant avec les modèles classiques, voire les surpassant, tout en restant dans des contraintes biologiques réalistes tirées de l’anatomie et de la physiologie.
Plusieurs chemins mènent au même sens du mouvement
La recherche a révélé que de simples différences dans la disposition spatiale des entrées peuvent générer une forte sélectivité de direction même lorsque leur timing est identique. Par exemple, disposer des cellules présynaptiques de sorte que leurs champs récepteurs augmentent progressivement en taille, ou tournent en orientation, le long de l’axe de mouvement préféré peut provoquer l’alignement temporel de leurs réponses pour un sens du mouvement mais pas pour l’autre. Les régions entourant la cellule qui suppriment sa réponse se sont aussi révélées puissantes : en faisant varier la force, la taille ou la vitesse de ces périphéries selon les entrées, le modèle pouvait définir des fenêtres temporelles précises où les signaux s’alignent uniquement lors du mouvement dans la direction préférée. Même des poids synaptiques inégaux, avec des filtres d’entrée identiques, permettaient aux circuits de devenir directionnels, bien que cette stratégie ait été moins performante que celles exploitant la structure des champs récepteurs.
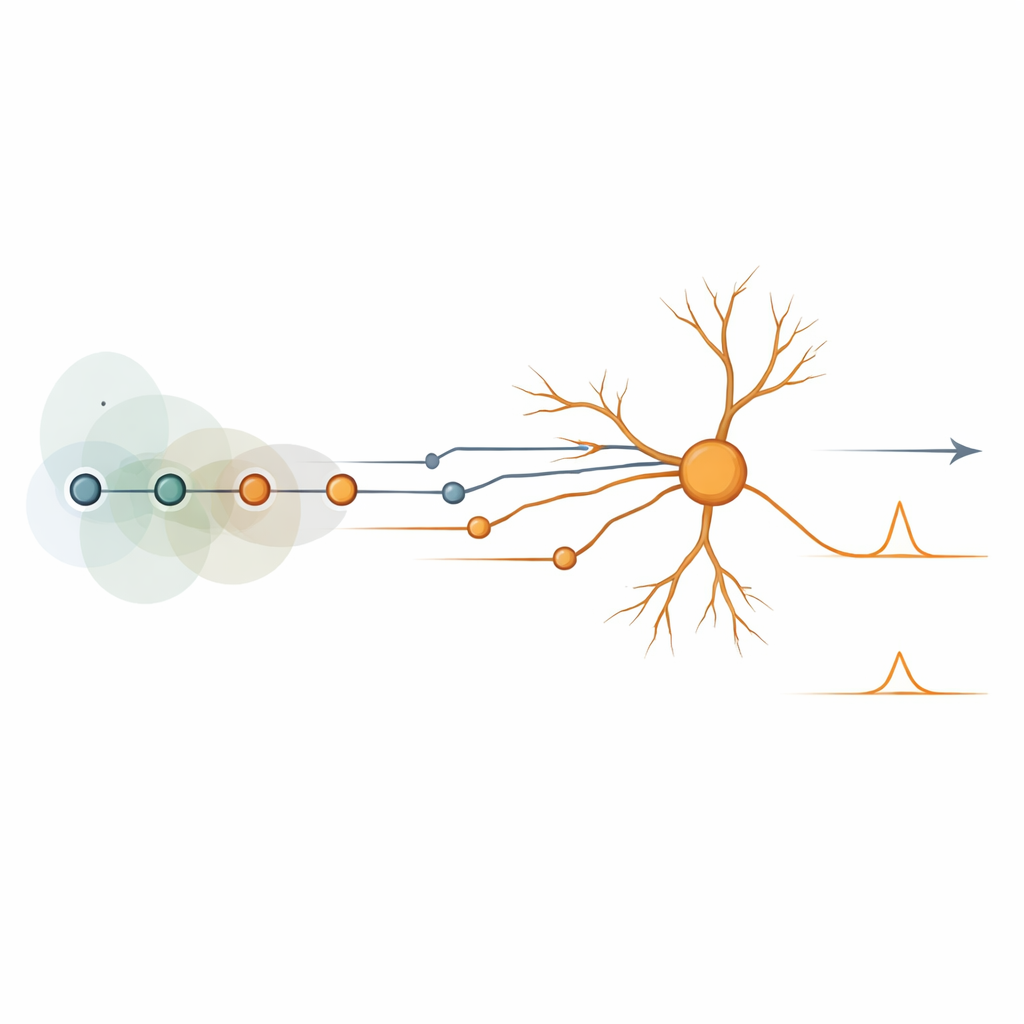
Blocs de construction cachés des circuits de mouvement Figure 2.
Malgré la variété déconcertante des circuits performants, leur comportement pouvait être distillé en un petit ensemble de « primitives computationnelles » — des blocs algorithmiques réutilisables. Certains correspondaient aux schémas classiques de retard-et-comparaison et d’inhibition. D’autres étaient nouveaux, comme des mécanismes qui s’appuient sur l’alignement des pics de nombreuses entrées dans le temps via des interactions spatiales et périphériques, ou des schémas où des pauses dans l’inhibition ou des changements d’amplitude de réponse véhiculent le signal directionnel. Lorsqu’on ajoutait des entrées inhibitrices, d’autres variantes apparaissaient, incluant une forme « anti » du modèle d’inhibition traditionnel et des configurations où l’inhibition chute brièvement au moment crucial pour laisser passer l’excitation. Ces primitives réapparaissaient non seulement dans des modèles détaillés de cellules ganglionnaires rétiniennes, mais aussi dans des neurones pyramidaux corticaux et dans des circuits jouets réduits à deux entrées, montrant qu’elles ne dépendent pas d’un type cellulaire ou d’un schéma de câblage particulier.
Détection du mouvement résiliente dans un monde bruyant
L’étude a également examiné la robustesse de ces mécanismes quand le mouvement est désordonné, comme dans la vie réelle où les vitesses varient de façon imprévisible. Les circuits qui s’appuyaient sur des changements de la taille des champs récepteurs d’entrée, en particulier dans leurs régions centrales, se sont révélés remarquablement robustes : ils continuaient à coder correctement la direction du mouvement même lorsque le stimulus s’accélérait et ralentissait de manière aléatoire. En revanche, les modèles qui dépendaient principalement de différences subtiles dans les propriétés périphériques ou d’autres relations temporelles finement accordées perdaient souvent leur sélectivité dans les mêmes conditions. Cela suggère que certaines des stratégies récemment découvertes peuvent être particulièrement adaptées à une perception du mouvement fiable dans des scènes naturelles.
Ce que cela signifie pour la compréhension du cerveau
En laissant l’apprentissage automatique explorer librement comment de simples circuits neuronaux pourraient détecter le mouvement, ce travail montre que le cerveau n’est pas limité à une ou deux astuces pour sentir le mouvement. Au contraire, de nombreuses dispositions de circuit distinctes se réduisent à une poignée d’idées computationnelles sous-jacentes qui peuvent être mises en œuvre avec des composants biologiques réalistes. Pour le grand public, la conclusion est que la détection du mouvement dans le cerveau est à la fois plus flexible et plus élégamment organisée qu’on ne le pensait : un petit ensemble d’opérations de base, réutilisées et recombinées, peut rendre compte d’une grande variété de cellules sensibles au mouvement à travers différentes régions cérébrales et espèces.
Citation: Poleg-Polsky, A. Machine learning discovers numerous new computational principles supporting elementary motion detection.
Nat Commun17, 3424 (2026). https://doi.org/10.1038/s41467-026-70288-4
Mots-clés: détection du mouvement, sélectivité de direction, circuits neuronaux, rétine et cortex, apprentissage automatique en neurosciences