Warum unser Gehirn sich für winzige Bewegungsverschiebungen interessiert
Ob Sie einen Ball fangen, ein vorbeifahrendes Auto bemerken oder einfach Blätter im Wind beobachten: Ihr visuelles System löst still und leise ein schwieriges Problem. Aus einem Durcheinander sich ändernder Lichtmuster auf der Netzhaut muss es ableiten, in welche Richtung sich Dinge bewegen. Jahrzehntelang stützten sich Forscher auf einige klassische Theorien, um zu erklären, wie Nervenzellen Bewegung erkennen. Diese Studie nutzt biologisch inspirierte Methoden des maschinellen Lernens, um eine kühner Frage zu stellen: Gibt es viel mehr Wege für einfache neuronale Schaltkreise, Bewegung zu erkennen, als wir bisher annahmen?
Klassische Ideen zur Bewegungswahrnehmung
Frühe Arbeiten zur Bewegungsvision schlugen vor, dass richtungssensitive Neuronen als kleine Zeitrechner fungieren. In einem traditionellen Modell werden Eingänge benachbarter Raumstellen verglichen, nachdem einer von ihnen verzögert wurde, sodass Signale, die in der bevorzugten Richtung wandern, gleichzeitig ankommen und sich summieren, während Bewegungen in die entgegengesetzte Richtung dies nicht tun. Ein anderes klassisches Modell stellt schnelle Erregung gegen verzögerte Hemmung aus der gegenüberliegenden Seite des rezeptiven Feldes eines Neurons, wodurch Antworten auf unerwünschte Richtungen ausgelöscht werden. Diese Mechanismen, obwohl einflussreich, konzentrieren sich hauptsächlich auf Zeitunterschiede und lassen offen, ob andere Schaltkreistricks denselben Zweck erfüllen könnten.
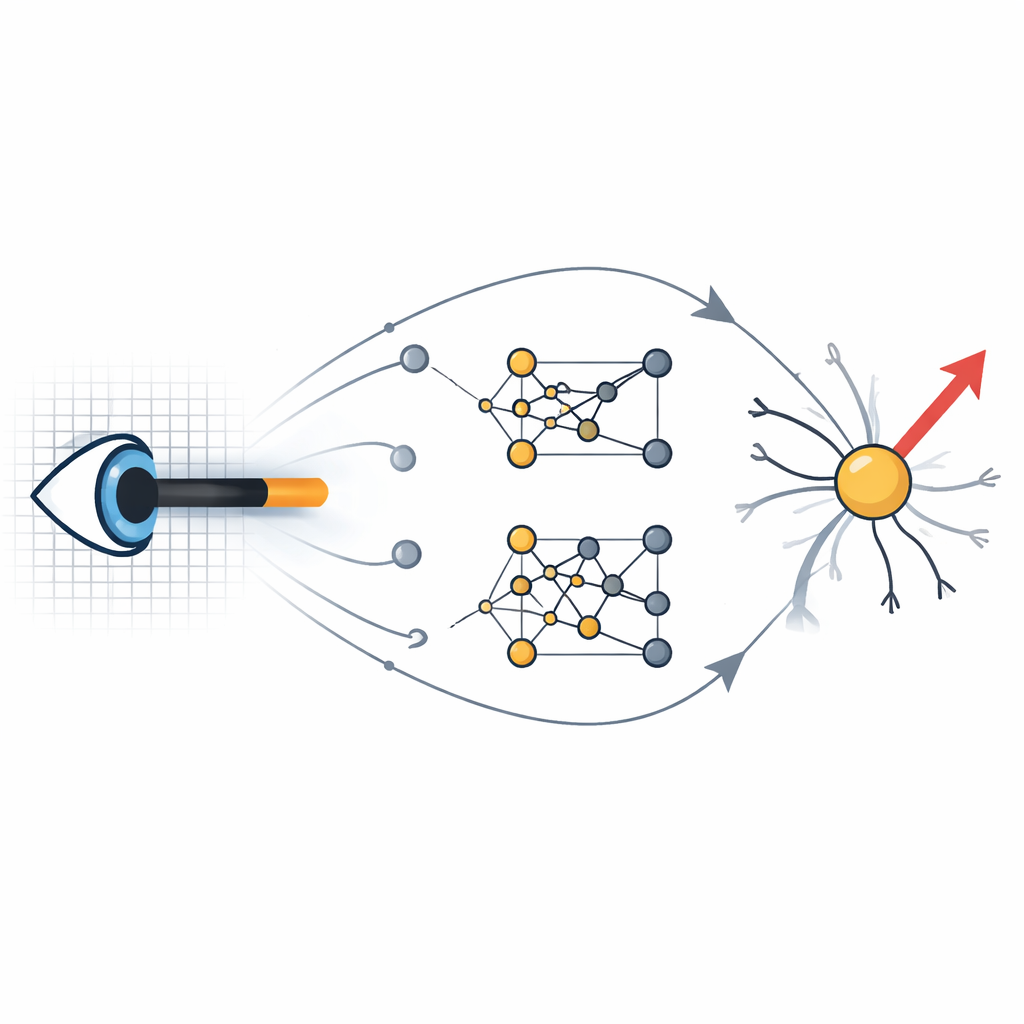
Die Evolution den Raum der Schaltkreise durchsuchen lassen Figure 1.
Um diese Frage zu untersuchen, entwickelte der Autor detaillierte Computermodelle von Neuronen in Retina und visuellem Kortex und setzte anschließend eine evolutionäre Suche auf. Ausgehend von zufälliger Verdrahtung und synaptischen Eigenschaften veränderte ein genetischer Algorithmus wiederholt Schaltkreisparameter und selektierte diejenigen Modelle, die die korrekte Richtung bewegter Balken und driftender Gitter am besten signalisierten. Wichtig war, dass die einzelnen Eingangszellen, die die Bewegungsdetektoren speisen, selbst keine Richtungssensitivität aufwiesen; nur ihre kollektive Anordnung und Dynamik konnte zu gerichteten Ausgaben führen. Diese automatisierte Erforschung förderte viele verschiedene Schaltkreislayouts zutage, die die Leistung klassischer Modelle erreichten oder übertrafen, und das alles innerhalb biologisch realistischer Beschränkungen, die aus Anatomie und Physiologie abgeleitet wurden.
Viele Wege zur selben Bewegungswahrnehmung
Die Suche zeigte, dass schon einfache Unterschiede in der räumlichen Anordnung der Eingänge starke Richtungsabstimmung erzeugen können, selbst wenn ihr Timing identisch ist. Beispielsweise kann das Anordnen präsynaptischer Zellen so, dass ihre rezeptiven Felder entlang der bevorzugten Bewegungsachse schrittweise größer werden oder sich in der Orientierung drehen, dazu führen, dass ihre Antworten in der Zeit für Bewegung in eine Richtung zusammenfallen, nicht aber in die andere. Umgebungsbereiche, die die Antwort einer Zelle unterdrücken, erwiesen sich ebenfalls als wirksam: Durch Variation von Stärke, Größe oder Geschwindigkeit dieser Umfelder über die Eingänge hinweg konnte das Modell präzise Zeitfenster ausschneiden, in denen Signale nur bei Bewegung in der bevorzugten Richtung zeitlich ausgerichtet sind. Selbst ungleiche synaptische Gewichte bei identischen Eingangsfiltern erlaubten es Schaltkreisen, richtungssensitiv zu werden, obwohl diese Strategie weniger erfolgreich war als solche, die die Struktur der rezeptiven Felder ausnutzten.
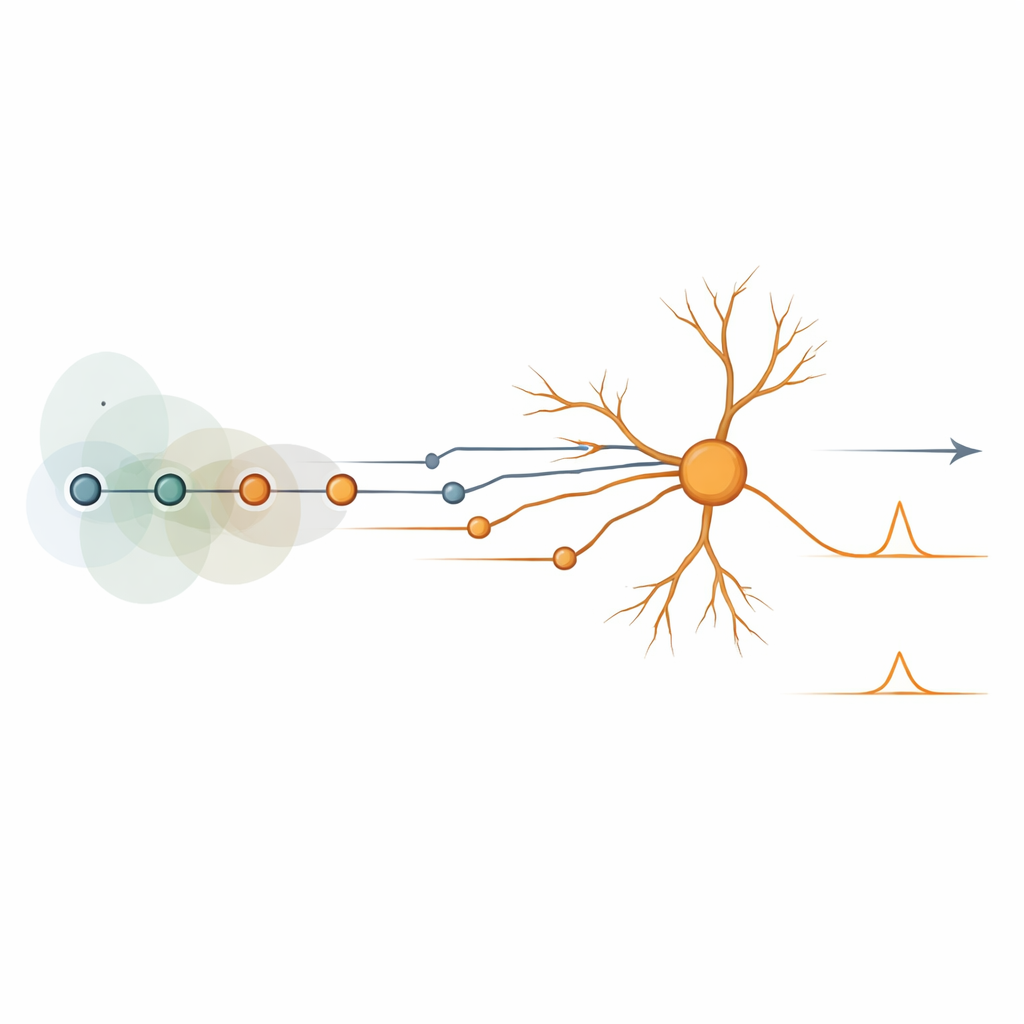
Verborgene Bausteine von Bewegungsschaltkreisen Figure 2.
Trotz der verwirrenden Vielfalt erfolgreicher Schaltkreise ließ sich ihr Verhalten auf eine kleine Menge „rechnerischer Primitive“ destillieren – wiederverwendbare algorithmische Bausteine. Einige entsprachen den klassischen Verzöger‑und‑Vergleich‑ sowie den hemmungsbasierten Schemata. Andere waren neu, etwa Mechanismen, die darauf beruhen, die Spitzen vieler Eingänge zeitlich mithilfe von Raum‑ und Umgebungsinteraktionen auszurichten, oder Schaltpläne, bei denen Pausen in der Hemmung oder Änderungen in der Antwortamplitude das Richtungssignal tragen. Mit Hinzufügen inhibitorischer Eingänge traten weitere Varianten auf, einschließlich einer „anti“‑Form des traditionellen Hemmungsmodells und Mustern, bei denen die Hemmung im entscheidenden Moment kurz absinkt, damit die Erregung durchscheint. Diese Primitive tauchten nicht nur in detaillierten Modellen der retinalen Ganglienzellen auf, sondern auch in kortikalen Pyramidenzellen und in reduzierten Zwei‑Eingangs‑Spielzeugschaltkreisen, was zeigt, dass sie nicht von einem bestimmten Zelltyp oder Verdrahtungsdiagramm abhängen.
Robuste Bewegungserkennung in einer lauten Welt
Die Studie untersuchte außerdem, wie diese Mechanismen sich verhalten, wenn Bewegung unordentlich ist, wie im echten Leben, wo Geschwindigkeiten unvorhersehbar variieren. Schaltkreise, die auf Änderungen der Größe der rezeptiven Felder der Eingänge setzten, insbesondere in ihren zentralen Regionen, erwiesen sich als bemerkenswert robust: Sie signalisierten die Bewegungsrichtung weiterhin genau, selbst wenn der Reiz zufällig beschleunigte und verlangsamte. Im Gegensatz dazu verloren Modelle, die hauptsächlich auf feinen Unterschieden in Umgebungsparametern oder anderen präzise abgestimmten zeitlichen Beziehungen beruhten, unter denselben Bedingungen eher ihre Selektivität. Das deutet darauf hin, dass einige der neu entdeckten Strategien besonders gut geeignet sein könnten für verlässliche Bewegungserkennung in natürlichen Szenen.
Was das für das Verständnis des Gehirns bedeutet
Indem maschinelles Lernen frei erforschen durfte, wie einfache neuronale Schaltkreise Bewegung erkennen könnten, zeigt diese Arbeit, dass das Gehirn nicht auf ein oder zwei Tricks zur Bewegungswahrnehmung beschränkt ist. Vielmehr lassen sich viele unterschiedliche Schaltkreisarrangements auf eine Handvoll zugrunde liegender rechnerischer Ideen zurückführen, die mit realistischen biologischen Komponenten umgesetzt werden können. Für nicht‑Fachleute lautet die Erkenntnis: Die Bewegungserkennung im Gehirn ist sowohl flexibler als auch eleganter organisiert als bisher gedacht: Eine kleine Menge Kernoperationen, wiederverwendet und neu kombiniert, kann eine breite Vielfalt bewegungssensitiver Zellen in verschiedenen Hirnregionen und Spezies erklären.
Zitation: Poleg-Polsky, A. Machine learning discovers numerous new computational principles supporting elementary motion detection.
Nat Commun17, 3424 (2026). https://doi.org/10.1038/s41467-026-70288-4
Schlüsselwörter: Bewegungserkennung, Richtungsspezifität, neuronale Schaltkreise, Retina und Kortex, Maschinelles Lernen in der Neurowissenschaft