Clear Sky Science · en
Dysregulated nuclear Lamin B1 in DYT1 dystonia thickens nuclear lamina and disrupts 14-3-3 proteins
When a Cell’s Inner Skeleton Goes Wrong
Childhood-onset DYT1 dystonia is a rare movement disorder that can twist muscles and contort posture, yet its root cause lies deep inside brain cells, at the boundary of the nucleus that holds our DNA. This study shows how a structural protein called Lamin B1, part of the cell’s “nuclear shell,” becomes mismanaged in DYT1 dystonia, warping the nucleus, blocking molecular traffic, and disrupting vital helper proteins that nerve cells need to grow and function properly.
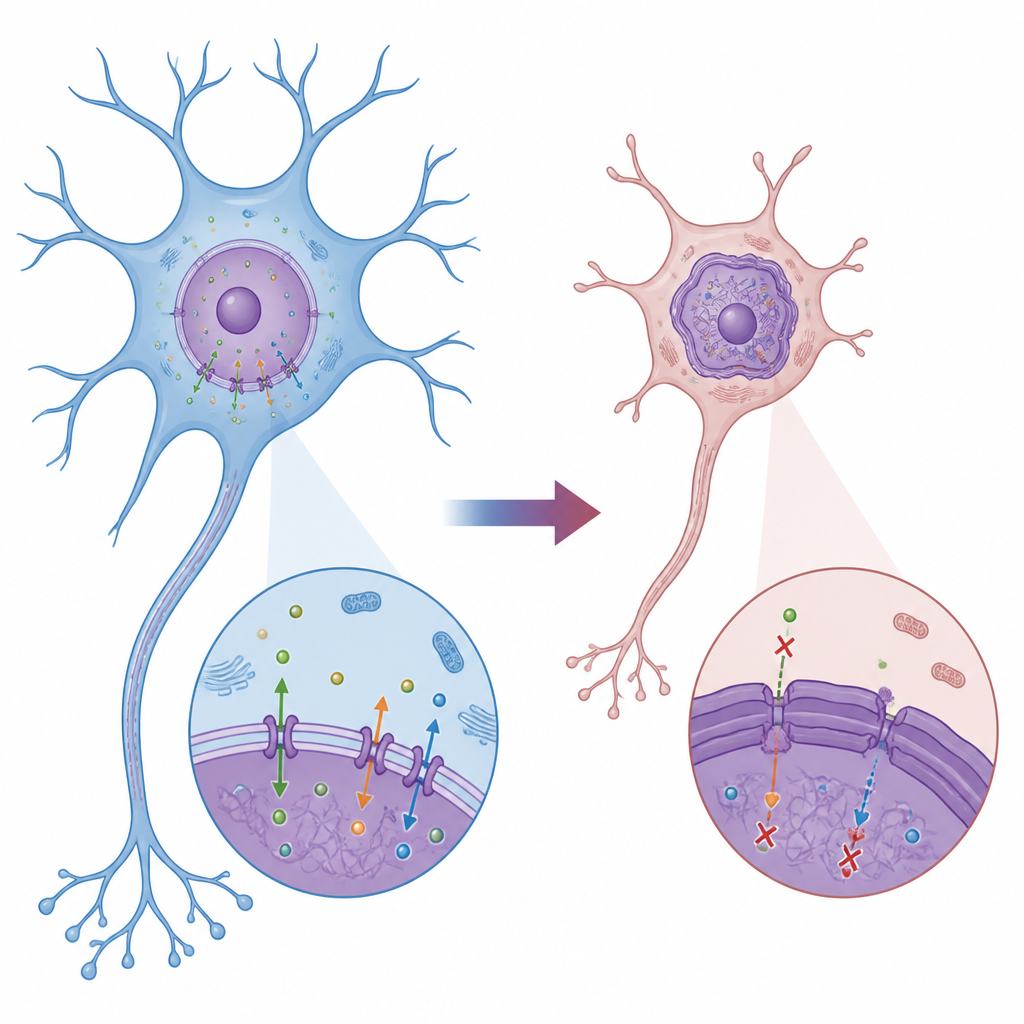
A Closer Look at a Childhood Movement Disorder
DYT1 dystonia usually begins in childhood or adolescence, a crucial time for brain circuits that control movement. Most cases are caused by a tiny three-letter deletion in the TOR1A gene, which encodes TorsinA, an enzyme that helps maintain the structure and transport systems around the nucleus. Earlier work in human neurons from patients hinted that Lamin B1, a key component of the nuclear “lamina” lining the inside of the nuclear membrane, was overabundant and misplaced. The current study set out to understand exactly how this Lamin B1 problem alters nuclear shape and function, and how those changes ripple out to damage developing neurons.
Warped Nuclei and Jammed Molecular Traffic
Using fibroblasts from DYT1 patients and age-matched healthy volunteers, the researchers found that cells with the DYT1 mutation had larger, misshapen nuclei and abnormally bright Lamin B1 staining, while a related protein, Lamin A/C, looked normal. Electron microscopy revealed a thickened dark band just under the inner nuclear membrane, showing that the nuclear lamina itself had become unusually bulky and rigid. Biochemical tests confirmed that some Lamin B1, which normally stays anchored at the nuclear edge, had leaked into the surrounding cytoplasm. These changes were linked to serious transport problems: fluorescent reporters and RNA probes showed that both proteins and messenger RNAs had trouble crossing the nuclear boundary, building up on the wrong side and indicating that the usual flow of information between nucleus and cytoplasm was impaired.
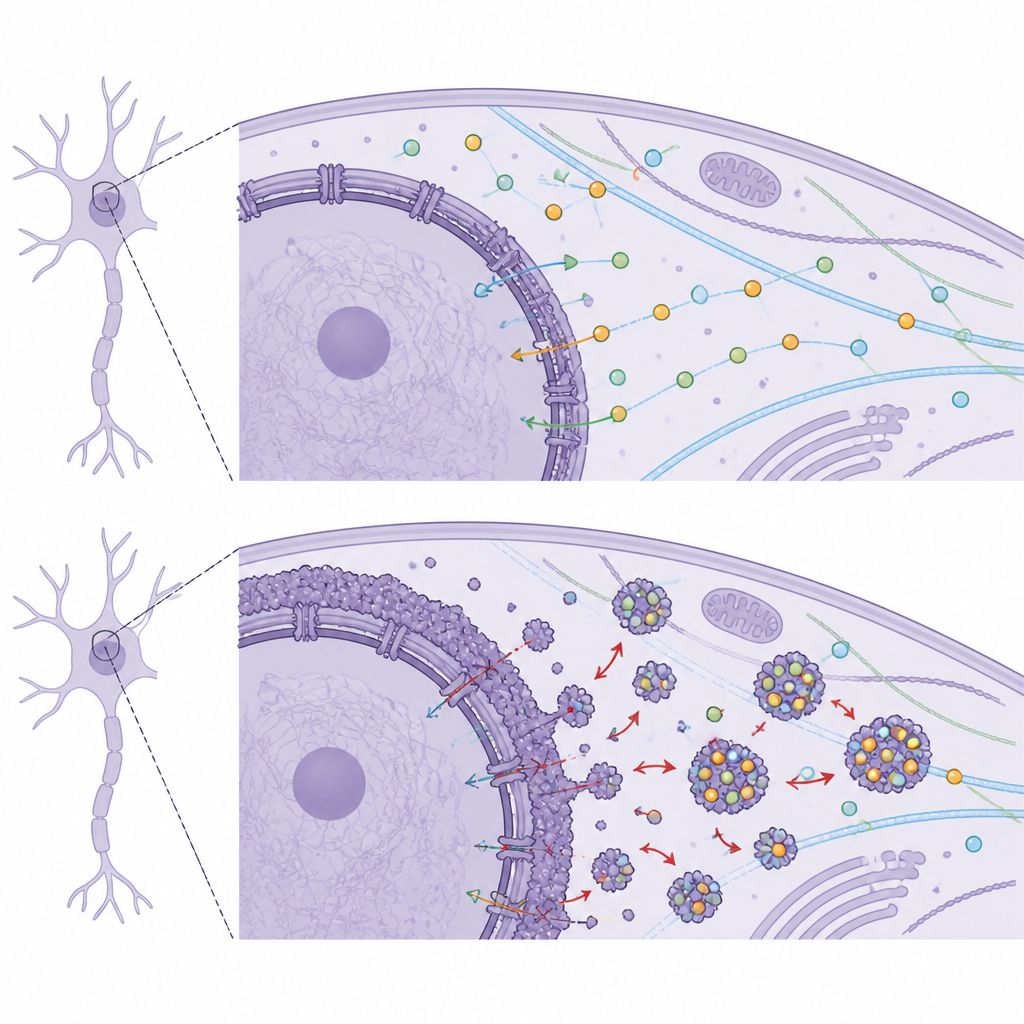
How Misplaced Lamin B1 Disrupts Neuron Machinery
To see what misplaced Lamin B1 might be “grabbing,” the team used human stem cell–derived motor neurons and a neuroblastoma-derived neuron model, forcing Lamin B1 into the cytoplasm and then fishing out its binding partners. Mass spectrometry revealed hundreds of interacting proteins involved in RNA processing, protein production, energy metabolism, cytoskeleton organization, and especially nucleocytoplasmic transport. Many of these partners are crucial for neuron-specific tasks such as axon growth, synapse formation, and signal transmission. A standout group among them were 14-3-3 proteins, a family of abundant chaperones that guide and stabilize other proteins during brain development and help control where they go inside the cell. Cytoplasmic Lamin B1 bound several 14-3-3 variants far more strongly than normal, suggesting it was sequestering these helpers away from their proper roles.
Helper Proteins that Shape Growing Neurons
The researchers next asked what happens when 14-3-3 proteins themselves are dialed up or down. In healthy stem cell–derived motor neurons, reducing two major 14-3-3 variants (known as beta and gamma) led to shorter neurites and fewer branches, along with lower levels of genes important for neuron maturation. In DYT1 neurons, several 14-3-3 genes were naturally reduced, and Lamin B1 was visibly mislocalized into neurites. When the team overexpressed 14-3-3 beta or gamma, DYT1 neurons grew longer, more branched processes and expressed higher levels of maturation markers. At the same time, Lamin B1 moved back toward the nucleus, its cytoplasmic buildup declined, and both protein and mRNA transport across the nuclear boundary improved, especially the import of proteins into the nucleus.
What This Means for Understanding and Treating Dystonia
In simple terms, this work links a flawed nuclear “scaffold” to stunted neuron growth in DYT1 dystonia. Too much Lamin B1, and Lamin B1 in the wrong place, thickens and stiffens the nuclear shell, warps the nucleus, jams molecular traffic, and ties up 14-3-3 helper proteins that neurons need to develop properly. By boosting 14-3-3 levels, the researchers could partially untangle this mess, restoring more normal Lamin B1 location, improving nuclear transport, and promoting healthier neurite growth. While these findings come from cell models rather than patients, they point to Lamin B1 and 14-3-3 proteins as attractive targets for future therapies aimed at protecting or restoring vulnerable motor neurons in DYT1 dystonia and potentially other neurological diseases that involve damaged nuclear architecture.
Citation: Duan, Y., Sepehrimanesh, M., Hosain, M.A. et al. Dysregulated nuclear Lamin B1 in DYT1 dystonia thickens nuclear lamina and disrupts 14-3-3 proteins. Cell Death Discov. 12, 245 (2026). https://doi.org/10.1038/s41420-026-03090-2
Keywords: DYT1 dystonia, Lamin B1, nuclear transport, 14-3-3 proteins, motor neurons